Entry
Reader's guide
Entries A-Z
Subject index
Heritability
The concept of heritability comes to us from population genetics. The idea was developed but not named by the geneticists Sewall Wright and Ronald Fisher. Both were theorists and applied their ideas to both agricultural and human psychological traits (i.e., intelligence). Wright developed the method of path analysis, which is now widely used in behavior genetics to compute heritability coefficients. These methods are very flexible and have been modified to incorporate modern molecular genetic findings. This entry outlines the purpose of the heritability formula, describes its various components, provides some examples of how the heritability of a trait is computed, and discusses some of the limitations of the concept (what it does not tell us).
The heritability coefficient indexes genetic influence on a quantitative trait (i.e., height, weight, intelligence) for a specific population. It tells us how much of the variance (a measure of the extent of differences between people) in a population is due to genetic factors. It is a population statistic (describes a population, just as the mean of a trait is descriptive of a population) and does not apply directly to individuals. It is useful to distinguish between “broad heritability” and “narrow heritability.”
Broad heritability tells us the proportion of total phenotypic (measurable) variance (σ2P) in a trait that is due to all genetic influences (σ2G). It can vary from sample to sample, environment to environment, and age to age. Thus the formula:
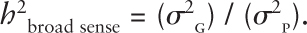
This formula should be read as broad heritability = all genetic variance/phenotypic variance. The term h2 is the symbol for heritability and is not to be interpreted to mean the value is to be squared.
We know from Mendelian theory that genes act in a number of ways. Many act additively: The more genes positive for the trait the more the trait expresses itself (height is a good example). The idea is that these additive genes, each of small effect, sum up to influence a trait. In addition, some genes dominate other genes (Mendelian dominance). This is a form of interaction (nonadditivity) between two genes at the same place (locus) on the paired human chromosomes. Another form of nonadditivity is epistasis, the interaction between genes at different loci on the chromosomes. Additive genes create similarity between relatives in a straightforward manner. Nonadditive effects work in a much more complex way and their effects are difficult to estimate with precision in human studies.
Narrow heritability includes only the proportion of variance because of genes that act in an additive manner. This is called the additive genetic variance (σ2A). Thus the formula:
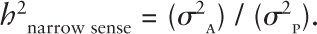
As in the previous example, this formula should be read as narrow heritability = additive genetic variance/phenotypic variance. Again the term h2 is the symbol for heritability and is not to be interpreted to mean the value is to be squared.
Environmental sources of variance are, of course, conceptually important. One such source is called shared environmental variance or common environmental variance (σ2C). This environmental influence is what has been thought to make twins and siblings similar (similar child rearing, same home, parents with the same educational background, etc.). For a long time psychologists thought that common environment was an extremely important source of twin and sibling similarity, but behavior genetic studies have shown that it is of minor importance for many traits measured in adulthood. For a number of traits (i.e., intelligence and social attitudes), shared environment is very important in the early years but much less so in adulthood. We can estimate the importance of shared environment in a number of ways, but two are particularly simple. Both use correlation coefficients. The correlation coefficient used in twin and adoption studies can run from zero to one and can, in the example below, be interpreted as a percentage of variance. In most other contexts, a correlation coefficient must be squared to interpret the results as a percentage. This point is widely misunderstood. Consider monozygotic twins reared together (MZT), often incorrectly called identical twins. They have all their genes in common. Consequently, the correlation between them on a trait such as intelligence reflects all the effects of genes (additive and nonadditive) plus the similarity because of being reared in the same family (shared environment). Monozygotic twins reared apart (MZA), if they were placed in adoptive homes randomly, would be similar only because they had all their genes in common (additive and nonadditive effects). Thus the MZA correlation would directly estimate the broad heritability. The difference between the two correlations (MZT – MZA) would estimate shared environmental influence. The IQ correlation between MZT twins, in adulthood, is about .86. The IQ correlation for MZA twins (almost all adults) is .74 (weighted mean of five studies). The difference suggests a shared environmental influence of .12. The second method of estimating shared environmental influence is to examine the similarity of unrelated individuals reared in the same home (URT). For childhood data the URT correlation is .26 (weighted mean of 14 studies). In adulthood the URT correlation is .04 (weighted mean of five studies). This method suggests a shared environmental influence of .04. Other methods yield estimates between these two figures and also show a strong age effect, with genetic influence increasing with age.
...
- Action and Motor Control
- Action and Bodily Movement
- Analogical Reasoning, Models of Development
- Anomalous Monism
- Anomalous Monism
- Anti-Individualism About Cognition
- Anti-Individualism About Cognition
- Attention and Emotions, Computational Perspectives
- Belief and Judgment
- Case-Based Reasoning, Computational Perspectives
- Causal Theories of Intentionality
- Causal Theories of Memory
- Computational Models of Emotion
- Computational Perspectives
- Conscious Thinking
- Conscious Thinking
- Consciousness and Embodiment
- Consciousness and Embodiment
- Consciousness and the Unconscious
- Deductive Reasoning
- Descriptive Thought
- Development
- Disorders and Pathology
- Disorders and Pathology
- Distributed Cognition
- Electrophysiological Studies of Mind
- Eliminative Materialism
- Eliminative Materialism
- Emergence
- Emergence
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Experimental Philosophy
- Explanatory Gap
- Explanatory Gap
- Extended Mind
- Face Recognition in Humans and Computers
- Facial Expressions, Computational Perspectives
- Facial Expressions, Computational Perspectives
- Freedom of Action
- Idealism
- Idealism
- Intelligence, Neural Basis
- Intentionality of Bodily Sensation
- Intentionality of Emotion
- Introspection
- Know-How, Philosophical Perspectives
- Know-How, Philosophical Perspectives
- Knowledge by Acquaintance
- Machine Speech Recognition
- Memory and Knowledge
- Mental Action
- Mental Causation
- Mental Causation
- Mind-Body Problem
- Modeling Causal Learning
- Moral Development
- Multimodal Conversational Systems
- Naïve Realism
- Natural Language Generation
- Neural Basis
- Object-Dependent Thought
- Personal Identity
- Personal Identity, Development of
- Philosophical Perspectives
- Philosophy of Action
- Physicalism
- Physicalism
- Psychological Research
- Realism and Instrumentalism
- Realism and Instrumentalism
- Reductive Physicalism
- Reductive Physicalism
- Relationships, Development of
- Representational Theory of Mind
- Self-Knowledge
- Semantic Memory, Computational Perspectives
- Sequential Memory, Computational Perspectives
- Serial Order Memory, Computational Perspectives
- Smell, Philosophical Perspectives
- Taste, Philosophical Perspectives
- Teleology
- Teleology
- Theory of Appearing
- Attention
- Aging, Memory, and Information Processing Speed
- Audition, Neural Basis
- Bilingual Language Processing
- Bilingualism, Cognitive Benefits of
- Black English Vernacular (Ebonics)
- Blindsight
- Borderline Personality Disorder
- Capgras Delusion
- Character and Personality, Philosophical Perspectives
- Collective Action
- Common Coding
- Computational Perspectives
- Distributed Cognition
- Emotion and Moral Judgment
- Emotion, Cultural Perspectives
- Emotional Recognition, Neuropsychology of
- Event Memory, Development
- Evolutionary Perspectives
- Experimental Philosophy
- Flynn Effect: Rising Intelligence Scores
- Gender Differences in Language and Language Use
- Gesture and Language Processing
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Heritability
- Heritage Language and Second Language Learning
- Intelligence and Working Memory
- Joint or Collective Intention
- Joint or Collective Intention
- Knowledge Acquisition in Development
- Mirror Neurons
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Multiple Intelligences Theory
- Narcissistic Personality Disorder
- Neural Basis
- Philosophical Perspectives
- Philosophy of Action
- Psychological Research
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Self-Knowledge
- Synesthesia
- Visual Imagery
- Word Learning
- Concepts and Categories
- Amnesia
- Analogical Reasoning, Models of Development
- Anxiety Disorders
- Character and Personality, Philosophical Perspectives
- Computational Perspectives
- Confabulation
- Desire
- Development
- Disjunctive Theory of Perception
- Disorders and Pathology
- Dissent, Effects on Group Decisions
- Distributed Cognition
- Dyslexia, Developmental
- Dyslexia, Phonological Processing in
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Exercise and the Brain
- Folk Psychology
- Group Decision Making
- Guilt
- Hearing, Philosophical Perspectives
- Heritage Language and Second Language Learning
- Human Classification Learning
- Innateness and Parameter Setting
- Intergroup Conflict
- Intergroup Conflict, Models of
- Knowledge Acquisition in Development
- Language Development
- Language Development, Overregulation in
- Moral Development
- Naïve Realism
- Neural Basis
- Philosophical Perspectives
- Political Psychology
- Prediction, Clinical Versus Actuarial
- Psychological Research
- Religion and Psychiatry
- Semantic Dementia
- Smell, Philosophical Perspectives
- Spatial Cognition, Development of
- Taste, Philosophical Perspectives
- Word Learning
- Consciousness
- Aphasia
- Attitude Change
- Attitude Change
- Attitudes and Behavior
- Attraction
- Attribution Theory
- Auditory Masking
- Behavioral Therapy
- Borderline Personality Disorder
- Cognitive Dissonance
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Common Coding
- Conduction Aphasia
- Consciousness, Comparative Perspectives
- Deductive Reasoning
- Depth Perception
- Desirable Difficulties Perspective on Learning
- Discrimination Learning, Training Methods
- Disorders and Pathology
- Dyslexia, Acquired
- Dyslexia, Developmental
- Dyslexia, Phonological Processing in
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Evolutionary Perspectives
- Exercise and the Brain
- Face Perception
- Facial Expressions, Emotional
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Group Decision Making
- Hearing
- Heritability
- Intergroup Conflict
- Intergroup Conflict, Models of
- Learning Styles
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Love
- McCollough Effect
- Meta-Analysis
- Metacognition and Education
- Motor Learning, Practical Aspects
- Multinomial Modeling
- Music Perception
- Neural Basis
- Optic Flow
- Perceptual Consciousness and Attention
- Perceptual Constancy
- Personal Identity, Development of
- Personality: Individual Versus Situation Debate
- Personality: Individual Versus Situation Debate
- Persuasion
- Philosophical Perspectives
- Placebo Effect
- Political Psychology
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Political Psychology
- Psychological Research
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Reaction Time
- Retrieval Practice (Testing) Effect
- Semantic Dementia
- Skill Learning, Enhancement of
- Social Cognition
- Social Loafing
- Spacing Effect, Practical Applications
- Speech Perception
- Stereopsis
- Subliminal Perception
- Synesthesia
- Time Perception
- Unconscious Perception
- Visual Imagery
- Visual Masking
- Visual Search
- Visuospatial Reasoning
- Wisdom of Crowds Effect
- Word Recognition, Auditory
- Word Recognition, Visual
- Working Memory, Evolution of
- Decision Making
- Amnesia
- Argument Mapping
- Behavioral Therapy
- Behaviorism
- Case-Based Reasoning, Computational Perspectives
- Computational Perspectives
- Desirable Difficulties Perspective on Learning
- Desire
- Desire
- Emotion and Psychopathology
- Emotion, Cerebral Lateralization
- Emotion, Psychophysiology of
- Emotional Recognition, Neuropsychology of
- Event Memory, Development
- Genes and Linguistic Tone
- Human Classification Learning
- Implicit Memory
- Learning Styles
- Memory, Neural Basis
- Metacognition and Education
- Music and the Evolution of Language
- Neural Basis
- Philosophical Perspectives
- Placebo Effect
- Practical Applications
- Psychological Research
- Reinforcement Learning, Psychological Perspectives
- Retrieval Practice (Testing) Effect
- Skill Learning, Enhancement of
- Spacing Effect
- Word Learning
- Disorders and Pathology
- Addiction
- Amnesia
- Analogical Mapping and Reasoning
- Anchoring
- Anosognosia
- Anxiety Disorders
- Aphasia
- Aphasia
- Apraxia
- Autism
- Automatic Behavior
- Autoscopic Phenomena
- Availability Heuristic
- Behavioral Therapy
- Borderline Personality Disorder
- Capgras Delusion
- Causal Theories of Memory
- Conduction Aphasia
- Conduction Aphasia
- Confabulation
- Delusions
- Desire
- Distributed Cognition
- Dyslexia, Acquired
- Dyslexia, Developmental
- Dyslexia, Phonological Processing in
- Emotion and Moral Judgment
- Emotion and Psychopathology
- Emotions and Consciousness
- Fregoli Delusion
- Hypochondria
- Intentionality of Emotion
- Legal Reasoning, Psychological Perspectives
- Memory and Knowledge
- Motivated Thinking
- Narcissistic Personality Disorder
- Objects of Memory
- Rationality of Emotion
- Religion and Psychiatry
- Representativeness Heuristic
- Schizophrenia
- Scientific Reasoning
- Semantic Dementia
- Spatial Cognition, Development of
- Thinking
- Two System Models of Reasoning
- Visual Imagery
- Visuospatial Reasoning
- Williams Syndrome
- Computational Perspectives
- Action and Bodily Movement
- Analogical Reasoning, Models of Development
- Anomalous Monism
- Anomalous Monism
- Anti-Individualism About Cognition
- Anti-Individualism About Cognition
- Attention and Emotions, Computational Perspectives
- Belief and Judgment
- Case-Based Reasoning, Computational Perspectives
- Causal Theories of Intentionality
- Causal Theories of Memory
- Computational Models of Emotion
- Computational Perspectives
- Conscious Thinking
- Conscious Thinking
- Consciousness and Embodiment
- Consciousness and Embodiment
- Consciousness and the Unconscious
- Deductive Reasoning
- Descriptive Thought
- Development
- Disorders and Pathology
- Disorders and Pathology
- Distributed Cognition
- Electrophysiological Studies of Mind
- Eliminative Materialism
- Eliminative Materialism
- Emergence
- Emergence
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Experimental Philosophy
- Explanatory Gap
- Explanatory Gap
- Extended Mind
- Face Recognition in Humans and Computers
- Facial Expressions, Computational Perspectives
- Facial Expressions, Computational Perspectives
- Freedom of Action
- Idealism
- Idealism
- Intelligence, Neural Basis
- Intentionality of Bodily Sensation
- Intentionality of Emotion
- Introspection
- Know-How, Philosophical Perspectives
- Know-How, Philosophical Perspectives
- Knowledge by Acquaintance
- Machine Speech Recognition
- Memory and Knowledge
- Mental Action
- Mental Causation
- Mental Causation
- Mind-Body Problem
- Modeling Causal Learning
- Moral Development
- Multimodal Conversational Systems
- Naïve Realism
- Natural Language Generation
- Neural Basis
- Object-Dependent Thought
- Personal Identity
- Personal Identity, Development of
- Philosophical Perspectives
- Philosophy of Action
- Physicalism
- Physicalism
- Psychological Research
- Realism and Instrumentalism
- Realism and Instrumentalism
- Reductive Physicalism
- Reductive Physicalism
- Relationships, Development of
- Representational Theory of Mind
- Self-Knowledge
- Semantic Memory, Computational Perspectives
- Sequential Memory, Computational Perspectives
- Serial Order Memory, Computational Perspectives
- Smell, Philosophical Perspectives
- Taste, Philosophical Perspectives
- Teleology
- Teleology
- Theory of Appearing
- Cultural Perspectives
- Aging, Memory, and Information Processing Speed
- Audition, Neural Basis
- Bilingual Language Processing
- Bilingualism, Cognitive Benefits of
- Black English Vernacular (Ebonics)
- Blindsight
- Borderline Personality Disorder
- Capgras Delusion
- Character and Personality, Philosophical Perspectives
- Collective Action
- Common Coding
- Computational Perspectives
- Distributed Cognition
- Emotion and Moral Judgment
- Emotion, Cultural Perspectives
- Emotional Recognition, Neuropsychology of
- Event Memory, Development
- Evolutionary Perspectives
- Experimental Philosophy
- Flynn Effect: Rising Intelligence Scores
- Gender Differences in Language and Language Use
- Gesture and Language Processing
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Heritability
- Heritage Language and Second Language Learning
- Intelligence and Working Memory
- Joint or Collective Intention
- Joint or Collective Intention
- Knowledge Acquisition in Development
- Mirror Neurons
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Multiple Intelligences Theory
- Narcissistic Personality Disorder
- Neural Basis
- Philosophical Perspectives
- Philosophy of Action
- Psychological Research
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Self-Knowledge
- Synesthesia
- Visual Imagery
- Word Learning
- Disorders and Pathology
- Amnesia
- Analogical Reasoning, Models of Development
- Anxiety Disorders
- Character and Personality, Philosophical Perspectives
- Computational Perspectives
- Confabulation
- Desire
- Development
- Disjunctive Theory of Perception
- Disorders and Pathology
- Dissent, Effects on Group Decisions
- Distributed Cognition
- Dyslexia, Developmental
- Dyslexia, Phonological Processing in
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Exercise and the Brain
- Folk Psychology
- Group Decision Making
- Guilt
- Hearing, Philosophical Perspectives
- Heritage Language and Second Language Learning
- Human Classification Learning
- Innateness and Parameter Setting
- Intergroup Conflict
- Intergroup Conflict, Models of
- Knowledge Acquisition in Development
- Language Development
- Language Development, Overregulation in
- Moral Development
- Naïve Realism
- Neural Basis
- Philosophical Perspectives
- Political Psychology
- Prediction, Clinical Versus Actuarial
- Psychological Research
- Religion and Psychiatry
- Semantic Dementia
- Smell, Philosophical Perspectives
- Spatial Cognition, Development of
- Taste, Philosophical Perspectives
- Word Learning
- Evolutionary Perspectives
- Aphasia
- Attitude Change
- Attitude Change
- Attitudes and Behavior
- Attraction
- Attribution Theory
- Auditory Masking
- Behavioral Therapy
- Borderline Personality Disorder
- Cognitive Dissonance
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Common Coding
- Conduction Aphasia
- Consciousness, Comparative Perspectives
- Deductive Reasoning
- Depth Perception
- Desirable Difficulties Perspective on Learning
- Discrimination Learning, Training Methods
- Disorders and Pathology
- Dyslexia, Acquired
- Dyslexia, Developmental
- Dyslexia, Phonological Processing in
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Evolutionary Perspectives
- Exercise and the Brain
- Face Perception
- Facial Expressions, Emotional
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Group Decision Making
- Hearing
- Heritability
- Intergroup Conflict
- Intergroup Conflict, Models of
- Learning Styles
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Love
- McCollough Effect
- Meta-Analysis
- Metacognition and Education
- Motor Learning, Practical Aspects
- Multinomial Modeling
- Music Perception
- Neural Basis
- Optic Flow
- Perceptual Consciousness and Attention
- Perceptual Constancy
- Personal Identity, Development of
- Personality: Individual Versus Situation Debate
- Personality: Individual Versus Situation Debate
- Persuasion
- Philosophical Perspectives
- Placebo Effect
- Political Psychology
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Political Psychology
- Psychological Research
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Reaction Time
- Retrieval Practice (Testing) Effect
- Semantic Dementia
- Skill Learning, Enhancement of
- Social Cognition
- Social Loafing
- Spacing Effect, Practical Applications
- Speech Perception
- Stereopsis
- Subliminal Perception
- Synesthesia
- Time Perception
- Unconscious Perception
- Visual Imagery
- Visual Masking
- Visual Search
- Visuospatial Reasoning
- Wisdom of Crowds Effect
- Word Recognition, Auditory
- Word Recognition, Visual
- Working Memory, Evolution of
- Neural Basis
- Amnesia
- Argument Mapping
- Behavioral Therapy
- Behaviorism
- Case-Based Reasoning, Computational Perspectives
- Computational Perspectives
- Desirable Difficulties Perspective on Learning
- Desire
- Desire
- Emotion and Psychopathology
- Emotion, Cerebral Lateralization
- Emotion, Psychophysiology of
- Emotional Recognition, Neuropsychology of
- Event Memory, Development
- Genes and Linguistic Tone
- Human Classification Learning
- Implicit Memory
- Learning Styles
- Memory, Neural Basis
- Metacognition and Education
- Music and the Evolution of Language
- Neural Basis
- Philosophical Perspectives
- Placebo Effect
- Practical Applications
- Psychological Research
- Reinforcement Learning, Psychological Perspectives
- Retrieval Practice (Testing) Effect
- Skill Learning, Enhancement of
- Spacing Effect
- Word Learning
- Philosophical Perspectives
- Addiction
- Amnesia
- Analogical Mapping and Reasoning
- Anchoring
- Anosognosia
- Anxiety Disorders
- Aphasia
- Aphasia
- Apraxia
- Autism
- Automatic Behavior
- Autoscopic Phenomena
- Availability Heuristic
- Behavioral Therapy
- Borderline Personality Disorder
- Capgras Delusion
- Causal Theories of Memory
- Conduction Aphasia
- Conduction Aphasia
- Confabulation
- Delusions
- Desire
- Distributed Cognition
- Dyslexia, Acquired
- Dyslexia, Developmental
- Dyslexia, Phonological Processing in
- Emotion and Moral Judgment
- Emotion and Psychopathology
- Emotions and Consciousness
- Fregoli Delusion
- Hypochondria
- Intentionality of Emotion
- Legal Reasoning, Psychological Perspectives
- Memory and Knowledge
- Motivated Thinking
- Narcissistic Personality Disorder
- Objects of Memory
- Rationality of Emotion
- Religion and Psychiatry
- Representativeness Heuristic
- Schizophrenia
- Scientific Reasoning
- Semantic Dementia
- Spatial Cognition, Development of
- Thinking
- Two System Models of Reasoning
- Visual Imagery
- Visuospatial Reasoning
- Williams Syndrome
- Psychological Research
- Affective Forecasting
- Anxiety Disorders
- Attention and Emotion
- Cognitive Dissonance
- Descriptions
- Descriptive Thought
- Disgust
- Embarrassment
- Emotion and Moral Judgment
- Emotion and Working Memory
- Emotion Regulation
- Emotion, Psychophysiology of
- Emotion, Structural Approaches
- Envy
- Exercise and the Brain
- Facial Expressions, Emotional
- Guilt
- Happiness
- Indexical Thought
- Intension and Extension
- Jealousy
- Love
- Metaphor
- Mnemonic Strategies
- Object-Dependent Thought
- Regret
- Religion and Psychiatry
- Resentment
- Retrieval Practice (Testing) Effect
- Spacing Effect, Practical Applications
- Unconscious Emotions, Psychological Perspectives
- Philosophical Perspectives
- Action and Bodily Movement
- Analogical Reasoning, Models of Development
- Anomalous Monism
- Anomalous Monism
- Anti-Individualism About Cognition
- Anti-Individualism About Cognition
- Attention and Emotions, Computational Perspectives
- Belief and Judgment
- Case-Based Reasoning, Computational Perspectives
- Causal Theories of Intentionality
- Causal Theories of Memory
- Computational Models of Emotion
- Computational Perspectives
- Conscious Thinking
- Conscious Thinking
- Consciousness and Embodiment
- Consciousness and Embodiment
- Consciousness and the Unconscious
- Deductive Reasoning
- Descriptive Thought
- Development
- Disorders and Pathology
- Disorders and Pathology
- Distributed Cognition
- Electrophysiological Studies of Mind
- Eliminative Materialism
- Eliminative Materialism
- Emergence
- Emergence
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Experimental Philosophy
- Explanatory Gap
- Explanatory Gap
- Extended Mind
- Face Recognition in Humans and Computers
- Facial Expressions, Computational Perspectives
- Facial Expressions, Computational Perspectives
- Freedom of Action
- Idealism
- Idealism
- Intelligence, Neural Basis
- Intentionality of Bodily Sensation
- Intentionality of Emotion
- Introspection
- Know-How, Philosophical Perspectives
- Know-How, Philosophical Perspectives
- Knowledge by Acquaintance
- Machine Speech Recognition
- Memory and Knowledge
- Mental Action
- Mental Causation
- Mental Causation
- Mind-Body Problem
- Modeling Causal Learning
- Moral Development
- Multimodal Conversational Systems
- Naïve Realism
- Natural Language Generation
- Neural Basis
- Object-Dependent Thought
- Personal Identity
- Personal Identity, Development of
- Philosophical Perspectives
- Philosophy of Action
- Physicalism
- Physicalism
- Psychological Research
- Realism and Instrumentalism
- Realism and Instrumentalism
- Reductive Physicalism
- Reductive Physicalism
- Relationships, Development of
- Representational Theory of Mind
- Self-Knowledge
- Semantic Memory, Computational Perspectives
- Sequential Memory, Computational Perspectives
- Serial Order Memory, Computational Perspectives
- Smell, Philosophical Perspectives
- Taste, Philosophical Perspectives
- Teleology
- Teleology
- Theory of Appearing
- Psychological Research
- Aging, Memory, and Information Processing Speed
- Audition, Neural Basis
- Bilingual Language Processing
- Bilingualism, Cognitive Benefits of
- Black English Vernacular (Ebonics)
- Blindsight
- Borderline Personality Disorder
- Capgras Delusion
- Character and Personality, Philosophical Perspectives
- Collective Action
- Common Coding
- Computational Perspectives
- Distributed Cognition
- Emotion and Moral Judgment
- Emotion, Cultural Perspectives
- Emotional Recognition, Neuropsychology of
- Event Memory, Development
- Evolutionary Perspectives
- Experimental Philosophy
- Flynn Effect: Rising Intelligence Scores
- Gender Differences in Language and Language Use
- Gesture and Language Processing
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Heritability
- Heritage Language and Second Language Learning
- Intelligence and Working Memory
- Joint or Collective Intention
- Joint or Collective Intention
- Knowledge Acquisition in Development
- Mirror Neurons
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Multiple Intelligences Theory
- Narcissistic Personality Disorder
- Neural Basis
- Philosophical Perspectives
- Philosophy of Action
- Psychological Research
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Self-Knowledge
- Synesthesia
- Visual Imagery
- Word Learning
- Heritability
- Action and Bodily Movement
- Analogical Reasoning, Models of Development
- Anomalous Monism
- Anomalous Monism
- Anti-Individualism About Cognition
- Anti-Individualism About Cognition
- Attention and Emotions, Computational Perspectives
- Belief and Judgment
- Case-Based Reasoning, Computational Perspectives
- Causal Theories of Intentionality
- Causal Theories of Memory
- Computational Models of Emotion
- Computational Perspectives
- Conscious Thinking
- Conscious Thinking
- Consciousness and Embodiment
- Consciousness and Embodiment
- Consciousness and the Unconscious
- Deductive Reasoning
- Descriptive Thought
- Development
- Disorders and Pathology
- Disorders and Pathology
- Distributed Cognition
- Electrophysiological Studies of Mind
- Eliminative Materialism
- Eliminative Materialism
- Emergence
- Emergence
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Experimental Philosophy
- Explanatory Gap
- Explanatory Gap
- Extended Mind
- Face Recognition in Humans and Computers
- Facial Expressions, Computational Perspectives
- Facial Expressions, Computational Perspectives
- Freedom of Action
- Idealism
- Idealism
- Intelligence, Neural Basis
- Intentionality of Bodily Sensation
- Intentionality of Emotion
- Introspection
- Know-How, Philosophical Perspectives
- Know-How, Philosophical Perspectives
- Knowledge by Acquaintance
- Machine Speech Recognition
- Memory and Knowledge
- Mental Action
- Mental Causation
- Mental Causation
- Mind-Body Problem
- Modeling Causal Learning
- Moral Development
- Multimodal Conversational Systems
- Naïve Realism
- Natural Language Generation
- Neural Basis
- Object-Dependent Thought
- Personal Identity
- Personal Identity, Development of
- Philosophical Perspectives
- Philosophy of Action
- Physicalism
- Physicalism
- Psychological Research
- Realism and Instrumentalism
- Realism and Instrumentalism
- Reductive Physicalism
- Reductive Physicalism
- Relationships, Development of
- Representational Theory of Mind
- Self-Knowledge
- Semantic Memory, Computational Perspectives
- Sequential Memory, Computational Perspectives
- Serial Order Memory, Computational Perspectives
- Smell, Philosophical Perspectives
- Taste, Philosophical Perspectives
- Teleology
- Teleology
- Theory of Appearing
- Cultural Perspectives
- Action and Bodily Movement
- Analogical Reasoning, Models of Development
- Anomalous Monism
- Anomalous Monism
- Anti-Individualism About Cognition
- Anti-Individualism About Cognition
- Attention and Emotions, Computational Perspectives
- Belief and Judgment
- Case-Based Reasoning, Computational Perspectives
- Causal Theories of Intentionality
- Causal Theories of Memory
- Computational Models of Emotion
- Computational Perspectives
- Conscious Thinking
- Conscious Thinking
- Consciousness and Embodiment
- Consciousness and Embodiment
- Consciousness and the Unconscious
- Deductive Reasoning
- Descriptive Thought
- Development
- Disorders and Pathology
- Disorders and Pathology
- Distributed Cognition
- Electrophysiological Studies of Mind
- Eliminative Materialism
- Eliminative Materialism
- Emergence
- Emergence
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Experimental Philosophy
- Explanatory Gap
- Explanatory Gap
- Extended Mind
- Face Recognition in Humans and Computers
- Facial Expressions, Computational Perspectives
- Facial Expressions, Computational Perspectives
- Freedom of Action
- Idealism
- Idealism
- Intelligence, Neural Basis
- Intentionality of Bodily Sensation
- Intentionality of Emotion
- Introspection
- Know-How, Philosophical Perspectives
- Know-How, Philosophical Perspectives
- Knowledge by Acquaintance
- Machine Speech Recognition
- Memory and Knowledge
- Mental Action
- Mental Causation
- Mental Causation
- Mind-Body Problem
- Modeling Causal Learning
- Moral Development
- Multimodal Conversational Systems
- Naïve Realism
- Natural Language Generation
- Neural Basis
- Object-Dependent Thought
- Personal Identity
- Personal Identity, Development of
- Philosophical Perspectives
- Philosophy of Action
- Physicalism
- Physicalism
- Psychological Research
- Realism and Instrumentalism
- Realism and Instrumentalism
- Reductive Physicalism
- Reductive Physicalism
- Relationships, Development of
- Representational Theory of Mind
- Self-Knowledge
- Semantic Memory, Computational Perspectives
- Sequential Memory, Computational Perspectives
- Serial Order Memory, Computational Perspectives
- Smell, Philosophical Perspectives
- Taste, Philosophical Perspectives
- Teleology
- Teleology
- Theory of Appearing
- Philosophical Perspectives
- Aging, Memory, and Information Processing Speed
- Audition, Neural Basis
- Bilingual Language Processing
- Bilingualism, Cognitive Benefits of
- Black English Vernacular (Ebonics)
- Blindsight
- Borderline Personality Disorder
- Capgras Delusion
- Character and Personality, Philosophical Perspectives
- Collective Action
- Common Coding
- Computational Perspectives
- Distributed Cognition
- Emotion and Moral Judgment
- Emotion, Cultural Perspectives
- Emotional Recognition, Neuropsychology of
- Event Memory, Development
- Evolutionary Perspectives
- Experimental Philosophy
- Flynn Effect: Rising Intelligence Scores
- Gender Differences in Language and Language Use
- Gesture and Language Processing
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Heritability
- Heritage Language and Second Language Learning
- Intelligence and Working Memory
- Joint or Collective Intention
- Joint or Collective Intention
- Knowledge Acquisition in Development
- Mirror Neurons
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Multiple Intelligences Theory
- Narcissistic Personality Disorder
- Neural Basis
- Philosophical Perspectives
- Philosophy of Action
- Psychological Research
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Self-Knowledge
- Synesthesia
- Visual Imagery
- Word Learning
- Psychological Research
- Amnesia
- Analogical Reasoning, Models of Development
- Anxiety Disorders
- Character and Personality, Philosophical Perspectives
- Computational Perspectives
- Confabulation
- Desire
- Development
- Disjunctive Theory of Perception
- Disorders and Pathology
- Dissent, Effects on Group Decisions
- Distributed Cognition
- Dyslexia, Developmental
- Dyslexia, Phonological Processing in
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Exercise and the Brain
- Folk Psychology
- Group Decision Making
- Guilt
- Hearing, Philosophical Perspectives
- Heritage Language and Second Language Learning
- Human Classification Learning
- Innateness and Parameter Setting
- Intergroup Conflict
- Intergroup Conflict, Models of
- Knowledge Acquisition in Development
- Language Development
- Language Development, Overregulation in
- Moral Development
- Naïve Realism
- Neural Basis
- Philosophical Perspectives
- Political Psychology
- Prediction, Clinical Versus Actuarial
- Psychological Research
- Religion and Psychiatry
- Semantic Dementia
- Smell, Philosophical Perspectives
- Spatial Cognition, Development of
- Taste, Philosophical Perspectives
- Word Learning
- Neural Basis
- Action and Bodily Movement
- Analogical Reasoning, Models of Development
- Anomalous Monism
- Anomalous Monism
- Anti-Individualism About Cognition
- Anti-Individualism About Cognition
- Attention and Emotions, Computational Perspectives
- Belief and Judgment
- Case-Based Reasoning, Computational Perspectives
- Causal Theories of Intentionality
- Causal Theories of Memory
- Computational Models of Emotion
- Computational Perspectives
- Conscious Thinking
- Conscious Thinking
- Consciousness and Embodiment
- Consciousness and Embodiment
- Consciousness and the Unconscious
- Deductive Reasoning
- Descriptive Thought
- Development
- Disorders and Pathology
- Disorders and Pathology
- Distributed Cognition
- Electrophysiological Studies of Mind
- Eliminative Materialism
- Eliminative Materialism
- Emergence
- Emergence
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Experimental Philosophy
- Explanatory Gap
- Explanatory Gap
- Extended Mind
- Face Recognition in Humans and Computers
- Facial Expressions, Computational Perspectives
- Facial Expressions, Computational Perspectives
- Freedom of Action
- Idealism
- Idealism
- Intelligence, Neural Basis
- Intentionality of Bodily Sensation
- Intentionality of Emotion
- Introspection
- Know-How, Philosophical Perspectives
- Know-How, Philosophical Perspectives
- Knowledge by Acquaintance
- Machine Speech Recognition
- Memory and Knowledge
- Mental Action
- Mental Causation
- Mental Causation
- Mind-Body Problem
- Modeling Causal Learning
- Moral Development
- Multimodal Conversational Systems
- Naïve Realism
- Natural Language Generation
- Neural Basis
- Object-Dependent Thought
- Personal Identity
- Personal Identity, Development of
- Philosophical Perspectives
- Philosophy of Action
- Physicalism
- Physicalism
- Psychological Research
- Realism and Instrumentalism
- Realism and Instrumentalism
- Reductive Physicalism
- Reductive Physicalism
- Relationships, Development of
- Representational Theory of Mind
- Self-Knowledge
- Semantic Memory, Computational Perspectives
- Sequential Memory, Computational Perspectives
- Serial Order Memory, Computational Perspectives
- Smell, Philosophical Perspectives
- Taste, Philosophical Perspectives
- Teleology
- Teleology
- Theory of Appearing
- Psychological Research
- Aging, Memory, and Information Processing Speed
- Audition, Neural Basis
- Bilingual Language Processing
- Bilingualism, Cognitive Benefits of
- Black English Vernacular (Ebonics)
- Blindsight
- Borderline Personality Disorder
- Capgras Delusion
- Character and Personality, Philosophical Perspectives
- Collective Action
- Common Coding
- Computational Perspectives
- Distributed Cognition
- Emotion and Moral Judgment
- Emotion, Cultural Perspectives
- Emotional Recognition, Neuropsychology of
- Event Memory, Development
- Evolutionary Perspectives
- Experimental Philosophy
- Flynn Effect: Rising Intelligence Scores
- Gender Differences in Language and Language Use
- Gesture and Language Processing
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Heritability
- Heritage Language and Second Language Learning
- Intelligence and Working Memory
- Joint or Collective Intention
- Joint or Collective Intention
- Knowledge Acquisition in Development
- Mirror Neurons
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Multiple Intelligences Theory
- Narcissistic Personality Disorder
- Neural Basis
- Philosophical Perspectives
- Philosophy of Action
- Psychological Research
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Self-Knowledge
- Synesthesia
- Visual Imagery
- Word Learning
- Computational Perspectives
- Action and Bodily Movement
- Analogical Reasoning, Models of Development
- Anomalous Monism
- Anomalous Monism
- Anti-Individualism About Cognition
- Anti-Individualism About Cognition
- Attention and Emotions, Computational Perspectives
- Belief and Judgment
- Case-Based Reasoning, Computational Perspectives
- Causal Theories of Intentionality
- Causal Theories of Memory
- Computational Models of Emotion
- Computational Perspectives
- Conscious Thinking
- Conscious Thinking
- Consciousness and Embodiment
- Consciousness and Embodiment
- Consciousness and the Unconscious
- Deductive Reasoning
- Descriptive Thought
- Development
- Disorders and Pathology
- Disorders and Pathology
- Distributed Cognition
- Electrophysiological Studies of Mind
- Eliminative Materialism
- Eliminative Materialism
- Emergence
- Emergence
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Experimental Philosophy
- Explanatory Gap
- Explanatory Gap
- Extended Mind
- Face Recognition in Humans and Computers
- Facial Expressions, Computational Perspectives
- Facial Expressions, Computational Perspectives
- Freedom of Action
- Idealism
- Idealism
- Intelligence, Neural Basis
- Intentionality of Bodily Sensation
- Intentionality of Emotion
- Introspection
- Know-How, Philosophical Perspectives
- Know-How, Philosophical Perspectives
- Knowledge by Acquaintance
- Machine Speech Recognition
- Memory and Knowledge
- Mental Action
- Mental Causation
- Mental Causation
- Mind-Body Problem
- Modeling Causal Learning
- Moral Development
- Multimodal Conversational Systems
- Naïve Realism
- Natural Language Generation
- Neural Basis
- Object-Dependent Thought
- Personal Identity
- Personal Identity, Development of
- Philosophical Perspectives
- Philosophy of Action
- Physicalism
- Physicalism
- Psychological Research
- Realism and Instrumentalism
- Realism and Instrumentalism
- Reductive Physicalism
- Reductive Physicalism
- Relationships, Development of
- Representational Theory of Mind
- Self-Knowledge
- Semantic Memory, Computational Perspectives
- Sequential Memory, Computational Perspectives
- Serial Order Memory, Computational Perspectives
- Smell, Philosophical Perspectives
- Taste, Philosophical Perspectives
- Teleology
- Teleology
- Theory of Appearing
- Cultural Perspectives
- Aging, Memory, and Information Processing Speed
- Audition, Neural Basis
- Bilingual Language Processing
- Bilingualism, Cognitive Benefits of
- Black English Vernacular (Ebonics)
- Blindsight
- Borderline Personality Disorder
- Capgras Delusion
- Character and Personality, Philosophical Perspectives
- Collective Action
- Common Coding
- Computational Perspectives
- Distributed Cognition
- Emotion and Moral Judgment
- Emotion, Cultural Perspectives
- Emotional Recognition, Neuropsychology of
- Event Memory, Development
- Evolutionary Perspectives
- Experimental Philosophy
- Flynn Effect: Rising Intelligence Scores
- Gender Differences in Language and Language Use
- Gesture and Language Processing
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Heritability
- Heritage Language and Second Language Learning
- Intelligence and Working Memory
- Joint or Collective Intention
- Joint or Collective Intention
- Knowledge Acquisition in Development
- Mirror Neurons
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Multiple Intelligences Theory
- Narcissistic Personality Disorder
- Neural Basis
- Philosophical Perspectives
- Philosophy of Action
- Psychological Research
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Self-Knowledge
- Synesthesia
- Visual Imagery
- Word Learning
- Development
- Amnesia
- Analogical Reasoning, Models of Development
- Anxiety Disorders
- Character and Personality, Philosophical Perspectives
- Computational Perspectives
- Confabulation
- Desire
- Development
- Disjunctive Theory of Perception
- Disorders and Pathology
- Dissent, Effects on Group Decisions
- Distributed Cognition
- Dyslexia, Developmental
- Dyslexia, Phonological Processing in
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Exercise and the Brain
- Folk Psychology
- Group Decision Making
- Guilt
- Hearing, Philosophical Perspectives
- Heritage Language and Second Language Learning
- Human Classification Learning
- Innateness and Parameter Setting
- Intergroup Conflict
- Intergroup Conflict, Models of
- Knowledge Acquisition in Development
- Language Development
- Language Development, Overregulation in
- Moral Development
- Naïve Realism
- Neural Basis
- Philosophical Perspectives
- Political Psychology
- Prediction, Clinical Versus Actuarial
- Psychological Research
- Religion and Psychiatry
- Semantic Dementia
- Smell, Philosophical Perspectives
- Spatial Cognition, Development of
- Taste, Philosophical Perspectives
- Word Learning
- Disorders and Pathology
- Aphasia
- Attitude Change
- Attitude Change
- Attitudes and Behavior
- Attraction
- Attribution Theory
- Auditory Masking
- Behavioral Therapy
- Borderline Personality Disorder
- Cognitive Dissonance
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Common Coding
- Conduction Aphasia
- Consciousness, Comparative Perspectives
- Deductive Reasoning
- Depth Perception
- Desirable Difficulties Perspective on Learning
- Discrimination Learning, Training Methods
- Disorders and Pathology
- Dyslexia, Acquired
- Dyslexia, Developmental
- Dyslexia, Phonological Processing in
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Evolutionary Perspectives
- Exercise and the Brain
- Face Perception
- Facial Expressions, Emotional
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Group Decision Making
- Hearing
- Heritability
- Intergroup Conflict
- Intergroup Conflict, Models of
- Learning Styles
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Love
- McCollough Effect
- Meta-Analysis
- Metacognition and Education
- Motor Learning, Practical Aspects
- Multinomial Modeling
- Music Perception
- Neural Basis
- Optic Flow
- Perceptual Consciousness and Attention
- Perceptual Constancy
- Personal Identity, Development of
- Personality: Individual Versus Situation Debate
- Personality: Individual Versus Situation Debate
- Persuasion
- Philosophical Perspectives
- Placebo Effect
- Political Psychology
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Political Psychology
- Psychological Research
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Reaction Time
- Retrieval Practice (Testing) Effect
- Semantic Dementia
- Skill Learning, Enhancement of
- Social Cognition
- Social Loafing
- Spacing Effect, Practical Applications
- Speech Perception
- Stereopsis
- Subliminal Perception
- Synesthesia
- Time Perception
- Unconscious Perception
- Visual Imagery
- Visual Masking
- Visual Search
- Visuospatial Reasoning
- Wisdom of Crowds Effect
- Word Recognition, Auditory
- Word Recognition, Visual
- Working Memory, Evolution of
- Evolutionary Perspectives
- Amnesia
- Argument Mapping
- Behavioral Therapy
- Behaviorism
- Case-Based Reasoning, Computational Perspectives
- Computational Perspectives
- Desirable Difficulties Perspective on Learning
- Desire
- Desire
- Emotion and Psychopathology
- Emotion, Cerebral Lateralization
- Emotion, Psychophysiology of
- Emotional Recognition, Neuropsychology of
- Event Memory, Development
- Genes and Linguistic Tone
- Human Classification Learning
- Implicit Memory
- Learning Styles
- Memory, Neural Basis
- Metacognition and Education
- Music and the Evolution of Language
- Neural Basis
- Philosophical Perspectives
- Placebo Effect
- Practical Applications
- Psychological Research
- Reinforcement Learning, Psychological Perspectives
- Retrieval Practice (Testing) Effect
- Skill Learning, Enhancement of
- Spacing Effect
- Word Learning
- Neural Basis
- Addiction
- Amnesia
- Analogical Mapping and Reasoning
- Anchoring
- Anosognosia
- Anxiety Disorders
- Aphasia
- Aphasia
- Apraxia
- Autism
- Automatic Behavior
- Autoscopic Phenomena
- Availability Heuristic
- Behavioral Therapy
- Borderline Personality Disorder
- Capgras Delusion
- Causal Theories of Memory
- Conduction Aphasia
- Conduction Aphasia
- Confabulation
- Delusions
- Desire
- Distributed Cognition
- Dyslexia, Acquired
- Dyslexia, Developmental
- Dyslexia, Phonological Processing in
- Emotion and Moral Judgment
- Emotion and Psychopathology
- Emotions and Consciousness
- Fregoli Delusion
- Hypochondria
- Intentionality of Emotion
- Legal Reasoning, Psychological Perspectives
- Memory and Knowledge
- Motivated Thinking
- Narcissistic Personality Disorder
- Objects of Memory
- Rationality of Emotion
- Religion and Psychiatry
- Representativeness Heuristic
- Schizophrenia
- Scientific Reasoning
- Semantic Dementia
- Spatial Cognition, Development of
- Thinking
- Two System Models of Reasoning
- Visual Imagery
- Visuospatial Reasoning
- Williams Syndrome
- Philosophical Perspectives
- Affective Forecasting
- Anxiety Disorders
- Attention and Emotion
- Cognitive Dissonance
- Descriptions
- Descriptive Thought
- Disgust
- Embarrassment
- Emotion and Moral Judgment
- Emotion and Working Memory
- Emotion Regulation
- Emotion, Psychophysiology of
- Emotion, Structural Approaches
- Envy
- Exercise and the Brain
- Facial Expressions, Emotional
- Guilt
- Happiness
- Indexical Thought
- Intension and Extension
- Jealousy
- Love
- Metaphor
- Mnemonic Strategies
- Object-Dependent Thought
- Regret
- Religion and Psychiatry
- Resentment
- Retrieval Practice (Testing) Effect
- Spacing Effect, Practical Applications
- Unconscious Emotions, Psychological Perspectives
- Practical Applications
- Aging, Memory, and Information Processing Speed
- Deception, Linguistic Cues to
- Divided Attention and Memory
- Emotion and Working Memory
- Event Memory, Development
- Exercise and the Brain
- Eyewitness Memory
- Implicit Memory
- Intelligence and Working Memory
- Lie Detection
- Memory Recall, Dynamics
- Memory, Interference With
- Rehearsal and Memory
- Retrieval Practice (Testing) Effect
- Semantic Memory
- Spacing Effect
- Visual Working Memory
- Working Memory
- Working Memory in Language Processing
- Psychological Research
- Bilingual Language Processing
- Bilingualism, Cognitive Benefits of
- Cohort Model of Auditory Word Recognition
- Compound Words, Processing of
- Concepts and Language
- Conceptual Combination
- Conversation and Dialogue
- Declarative/Procedural Model of Language
- Discourse Processing, Models of
- Disfluencies: Comprehension Processes
- Eye Movements During Reading
- Frequency Effects in Word Recognition
- Gesture and Language Processing
- Inferences in Language Comprehension
- Inner Speech
- Language Development
- Language Production, Agreement in
- Language Production, Incremental Processing in
- Lie Detection
- Metaphor
- Perspective Taking in Language Processing
- Planning in Language Production
- Production of Language
- Prosody in Production
- Speech Perception
- Syntactic Production, Agreement in
- Word Learning
- Word Recognition, Auditory
- Word Recognition, Visual
- Working Memory in Language Processing
- Computational Perspectives
- Action and Bodily Movement
- Analogical Reasoning, Models of Development
- Anomalous Monism
- Anomalous Monism
- Anti-Individualism About Cognition
- Anti-Individualism About Cognition
- Attention and Emotions, Computational Perspectives
- Belief and Judgment
- Case-Based Reasoning, Computational Perspectives
- Causal Theories of Intentionality
- Causal Theories of Memory
- Computational Models of Emotion
- Computational Perspectives
- Conscious Thinking
- Conscious Thinking
- Consciousness and Embodiment
- Consciousness and Embodiment
- Consciousness and the Unconscious
- Deductive Reasoning
- Descriptive Thought
- Development
- Disorders and Pathology
- Disorders and Pathology
- Distributed Cognition
- Electrophysiological Studies of Mind
- Eliminative Materialism
- Eliminative Materialism
- Emergence
- Emergence
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Experimental Philosophy
- Explanatory Gap
- Explanatory Gap
- Extended Mind
- Face Recognition in Humans and Computers
- Facial Expressions, Computational Perspectives
- Facial Expressions, Computational Perspectives
- Freedom of Action
- Idealism
- Idealism
- Intelligence, Neural Basis
- Intentionality of Bodily Sensation
- Intentionality of Emotion
- Introspection
- Know-How, Philosophical Perspectives
- Know-How, Philosophical Perspectives
- Knowledge by Acquaintance
- Machine Speech Recognition
- Memory and Knowledge
- Mental Action
- Mental Causation
- Mental Causation
- Mind-Body Problem
- Modeling Causal Learning
- Moral Development
- Multimodal Conversational Systems
- Naïve Realism
- Natural Language Generation
- Neural Basis
- Object-Dependent Thought
- Personal Identity
- Personal Identity, Development of
- Philosophical Perspectives
- Philosophy of Action
- Physicalism
- Physicalism
- Psychological Research
- Realism and Instrumentalism
- Realism and Instrumentalism
- Reductive Physicalism
- Reductive Physicalism
- Relationships, Development of
- Representational Theory of Mind
- Self-Knowledge
- Semantic Memory, Computational Perspectives
- Sequential Memory, Computational Perspectives
- Serial Order Memory, Computational Perspectives
- Smell, Philosophical Perspectives
- Taste, Philosophical Perspectives
- Teleology
- Teleology
- Theory of Appearing
- Development
- Aging, Memory, and Information Processing Speed
- Audition, Neural Basis
- Bilingual Language Processing
- Bilingualism, Cognitive Benefits of
- Black English Vernacular (Ebonics)
- Blindsight
- Borderline Personality Disorder
- Capgras Delusion
- Character and Personality, Philosophical Perspectives
- Collective Action
- Common Coding
- Computational Perspectives
- Distributed Cognition
- Emotion and Moral Judgment
- Emotion, Cultural Perspectives
- Emotional Recognition, Neuropsychology of
- Event Memory, Development
- Evolutionary Perspectives
- Experimental Philosophy
- Flynn Effect: Rising Intelligence Scores
- Gender Differences in Language and Language Use
- Gesture and Language Processing
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Heritability
- Heritage Language and Second Language Learning
- Intelligence and Working Memory
- Joint or Collective Intention
- Joint or Collective Intention
- Knowledge Acquisition in Development
- Mirror Neurons
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Multiple Intelligences Theory
- Narcissistic Personality Disorder
- Neural Basis
- Philosophical Perspectives
- Philosophy of Action
- Psychological Research
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Self-Knowledge
- Synesthesia
- Visual Imagery
- Word Learning
- Neural Basis
- Amnesia
- Analogical Reasoning, Models of Development
- Anxiety Disorders
- Character and Personality, Philosophical Perspectives
- Computational Perspectives
- Confabulation
- Desire
- Development
- Disjunctive Theory of Perception
- Disorders and Pathology
- Dissent, Effects on Group Decisions
- Distributed Cognition
- Dyslexia, Developmental
- Dyslexia, Phonological Processing in
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Exercise and the Brain
- Folk Psychology
- Group Decision Making
- Guilt
- Hearing, Philosophical Perspectives
- Heritage Language and Second Language Learning
- Human Classification Learning
- Innateness and Parameter Setting
- Intergroup Conflict
- Intergroup Conflict, Models of
- Knowledge Acquisition in Development
- Language Development
- Language Development, Overregulation in
- Moral Development
- Naïve Realism
- Neural Basis
- Philosophical Perspectives
- Political Psychology
- Prediction, Clinical Versus Actuarial
- Psychological Research
- Religion and Psychiatry
- Semantic Dementia
- Smell, Philosophical Perspectives
- Spatial Cognition, Development of
- Taste, Philosophical Perspectives
- Word Learning
- Practical Applications
- Aphasia
- Attitude Change
- Attitude Change
- Attitudes and Behavior
- Attraction
- Attribution Theory
- Auditory Masking
- Behavioral Therapy
- Borderline Personality Disorder
- Cognitive Dissonance
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Common Coding
- Conduction Aphasia
- Consciousness, Comparative Perspectives
- Deductive Reasoning
- Depth Perception
- Desirable Difficulties Perspective on Learning
- Discrimination Learning, Training Methods
- Disorders and Pathology
- Dyslexia, Acquired
- Dyslexia, Developmental
- Dyslexia, Phonological Processing in
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Evolutionary Perspectives
- Exercise and the Brain
- Face Perception
- Facial Expressions, Emotional
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Group Decision Making
- Hearing
- Heritability
- Intergroup Conflict
- Intergroup Conflict, Models of
- Learning Styles
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Love
- McCollough Effect
- Meta-Analysis
- Metacognition and Education
- Motor Learning, Practical Aspects
- Multinomial Modeling
- Music Perception
- Neural Basis
- Optic Flow
- Perceptual Consciousness and Attention
- Perceptual Constancy
- Personal Identity, Development of
- Personality: Individual Versus Situation Debate
- Personality: Individual Versus Situation Debate
- Persuasion
- Philosophical Perspectives
- Placebo Effect
- Political Psychology
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Political Psychology
- Psychological Research
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Reaction Time
- Retrieval Practice (Testing) Effect
- Semantic Dementia
- Skill Learning, Enhancement of
- Social Cognition
- Social Loafing
- Spacing Effect, Practical Applications
- Speech Perception
- Stereopsis
- Subliminal Perception
- Synesthesia
- Time Perception
- Unconscious Perception
- Visual Imagery
- Visual Masking
- Visual Search
- Visuospatial Reasoning
- Wisdom of Crowds Effect
- Word Recognition, Auditory
- Word Recognition, Visual
- Working Memory, Evolution of
- Psychological Research
- Amnesia
- Argument Mapping
- Behavioral Therapy
- Behaviorism
- Case-Based Reasoning, Computational Perspectives
- Computational Perspectives
- Desirable Difficulties Perspective on Learning
- Desire
- Desire
- Emotion and Psychopathology
- Emotion, Cerebral Lateralization
- Emotion, Psychophysiology of
- Emotional Recognition, Neuropsychology of
- Event Memory, Development
- Genes and Linguistic Tone
- Human Classification Learning
- Implicit Memory
- Learning Styles
- Memory, Neural Basis
- Metacognition and Education
- Music and the Evolution of Language
- Neural Basis
- Philosophical Perspectives
- Placebo Effect
- Practical Applications
- Psychological Research
- Reinforcement Learning, Psychological Perspectives
- Retrieval Practice (Testing) Effect
- Skill Learning, Enhancement of
- Spacing Effect
- Word Learning
- Computational Perspectives
- Action and Bodily Movement
- Analogical Reasoning, Models of Development
- Anomalous Monism
- Anomalous Monism
- Anti-Individualism About Cognition
- Anti-Individualism About Cognition
- Attention and Emotions, Computational Perspectives
- Belief and Judgment
- Case-Based Reasoning, Computational Perspectives
- Causal Theories of Intentionality
- Causal Theories of Memory
- Computational Models of Emotion
- Computational Perspectives
- Conscious Thinking
- Conscious Thinking
- Consciousness and Embodiment
- Consciousness and Embodiment
- Consciousness and the Unconscious
- Deductive Reasoning
- Descriptive Thought
- Development
- Disorders and Pathology
- Disorders and Pathology
- Distributed Cognition
- Electrophysiological Studies of Mind
- Eliminative Materialism
- Eliminative Materialism
- Emergence
- Emergence
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Experimental Philosophy
- Explanatory Gap
- Explanatory Gap
- Extended Mind
- Face Recognition in Humans and Computers
- Facial Expressions, Computational Perspectives
- Facial Expressions, Computational Perspectives
- Freedom of Action
- Idealism
- Idealism
- Intelligence, Neural Basis
- Intentionality of Bodily Sensation
- Intentionality of Emotion
- Introspection
- Know-How, Philosophical Perspectives
- Know-How, Philosophical Perspectives
- Knowledge by Acquaintance
- Machine Speech Recognition
- Memory and Knowledge
- Mental Action
- Mental Causation
- Mental Causation
- Mind-Body Problem
- Modeling Causal Learning
- Moral Development
- Multimodal Conversational Systems
- Naïve Realism
- Natural Language Generation
- Neural Basis
- Object-Dependent Thought
- Personal Identity
- Personal Identity, Development of
- Philosophical Perspectives
- Philosophy of Action
- Physicalism
- Physicalism
- Psychological Research
- Realism and Instrumentalism
- Realism and Instrumentalism
- Reductive Physicalism
- Reductive Physicalism
- Relationships, Development of
- Representational Theory of Mind
- Self-Knowledge
- Semantic Memory, Computational Perspectives
- Sequential Memory, Computational Perspectives
- Serial Order Memory, Computational Perspectives
- Smell, Philosophical Perspectives
- Taste, Philosophical Perspectives
- Teleology
- Teleology
- Theory of Appearing
- Development
- Aging, Memory, and Information Processing Speed
- Audition, Neural Basis
- Bilingual Language Processing
- Bilingualism, Cognitive Benefits of
- Black English Vernacular (Ebonics)
- Blindsight
- Borderline Personality Disorder
- Capgras Delusion
- Character and Personality, Philosophical Perspectives
- Collective Action
- Common Coding
- Computational Perspectives
- Distributed Cognition
- Emotion and Moral Judgment
- Emotion, Cultural Perspectives
- Emotional Recognition, Neuropsychology of
- Event Memory, Development
- Evolutionary Perspectives
- Experimental Philosophy
- Flynn Effect: Rising Intelligence Scores
- Gender Differences in Language and Language Use
- Gesture and Language Processing
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Heritability
- Heritage Language and Second Language Learning
- Intelligence and Working Memory
- Joint or Collective Intention
- Joint or Collective Intention
- Knowledge Acquisition in Development
- Mirror Neurons
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Multiple Intelligences Theory
- Narcissistic Personality Disorder
- Neural Basis
- Philosophical Perspectives
- Philosophy of Action
- Psychological Research
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Self-Knowledge
- Synesthesia
- Visual Imagery
- Word Learning
- Disorders and Pathology
- Amnesia
- Analogical Reasoning, Models of Development
- Anxiety Disorders
- Character and Personality, Philosophical Perspectives
- Computational Perspectives
- Confabulation
- Desire
- Development
- Disjunctive Theory of Perception
- Disorders and Pathology
- Dissent, Effects on Group Decisions
- Distributed Cognition
- Dyslexia, Developmental
- Dyslexia, Phonological Processing in
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Exercise and the Brain
- Folk Psychology
- Group Decision Making
- Guilt
- Hearing, Philosophical Perspectives
- Heritage Language and Second Language Learning
- Human Classification Learning
- Innateness and Parameter Setting
- Intergroup Conflict
- Intergroup Conflict, Models of
- Knowledge Acquisition in Development
- Language Development
- Language Development, Overregulation in
- Moral Development
- Naïve Realism
- Neural Basis
- Philosophical Perspectives
- Political Psychology
- Prediction, Clinical Versus Actuarial
- Psychological Research
- Religion and Psychiatry
- Semantic Dementia
- Smell, Philosophical Perspectives
- Spatial Cognition, Development of
- Taste, Philosophical Perspectives
- Word Learning
- Evolutionary Perspectives
- Aphasia
- Attitude Change
- Attitude Change
- Attitudes and Behavior
- Attraction
- Attribution Theory
- Auditory Masking
- Behavioral Therapy
- Borderline Personality Disorder
- Cognitive Dissonance
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Common Coding
- Conduction Aphasia
- Consciousness, Comparative Perspectives
- Deductive Reasoning
- Depth Perception
- Desirable Difficulties Perspective on Learning
- Discrimination Learning, Training Methods
- Disorders and Pathology
- Dyslexia, Acquired
- Dyslexia, Developmental
- Dyslexia, Phonological Processing in
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Evolutionary Perspectives
- Exercise and the Brain
- Face Perception
- Facial Expressions, Emotional
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Group Decision Making
- Hearing
- Heritability
- Intergroup Conflict
- Intergroup Conflict, Models of
- Learning Styles
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Love
- McCollough Effect
- Meta-Analysis
- Metacognition and Education
- Motor Learning, Practical Aspects
- Multinomial Modeling
- Music Perception
- Neural Basis
- Optic Flow
- Perceptual Consciousness and Attention
- Perceptual Constancy
- Personal Identity, Development of
- Personality: Individual Versus Situation Debate
- Personality: Individual Versus Situation Debate
- Persuasion
- Philosophical Perspectives
- Placebo Effect
- Political Psychology
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Political Psychology
- Psychological Research
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Reaction Time
- Retrieval Practice (Testing) Effect
- Semantic Dementia
- Skill Learning, Enhancement of
- Social Cognition
- Social Loafing
- Spacing Effect, Practical Applications
- Speech Perception
- Stereopsis
- Subliminal Perception
- Synesthesia
- Time Perception
- Unconscious Perception
- Visual Imagery
- Visual Masking
- Visual Search
- Visuospatial Reasoning
- Wisdom of Crowds Effect
- Word Recognition, Auditory
- Word Recognition, Visual
- Working Memory, Evolution of
- Neural Basis
- Amnesia
- Argument Mapping
- Behavioral Therapy
- Behaviorism
- Case-Based Reasoning, Computational Perspectives
- Computational Perspectives
- Desirable Difficulties Perspective on Learning
- Desire
- Desire
- Emotion and Psychopathology
- Emotion, Cerebral Lateralization
- Emotion, Psychophysiology of
- Emotional Recognition, Neuropsychology of
- Event Memory, Development
- Genes and Linguistic Tone
- Human Classification Learning
- Implicit Memory
- Learning Styles
- Memory, Neural Basis
- Metacognition and Education
- Music and the Evolution of Language
- Neural Basis
- Philosophical Perspectives
- Placebo Effect
- Practical Applications
- Psychological Research
- Reinforcement Learning, Psychological Perspectives
- Retrieval Practice (Testing) Effect
- Skill Learning, Enhancement of
- Spacing Effect
- Word Learning
- Philosophical Perspectives
- Addiction
- Amnesia
- Analogical Mapping and Reasoning
- Anchoring
- Anosognosia
- Anxiety Disorders
- Aphasia
- Aphasia
- Apraxia
- Autism
- Automatic Behavior
- Autoscopic Phenomena
- Availability Heuristic
- Behavioral Therapy
- Borderline Personality Disorder
- Capgras Delusion
- Causal Theories of Memory
- Conduction Aphasia
- Conduction Aphasia
- Confabulation
- Delusions
- Desire
- Distributed Cognition
- Dyslexia, Acquired
- Dyslexia, Developmental
- Dyslexia, Phonological Processing in
- Emotion and Moral Judgment
- Emotion and Psychopathology
- Emotions and Consciousness
- Fregoli Delusion
- Hypochondria
- Intentionality of Emotion
- Legal Reasoning, Psychological Perspectives
- Memory and Knowledge
- Motivated Thinking
- Narcissistic Personality Disorder
- Objects of Memory
- Rationality of Emotion
- Religion and Psychiatry
- Representativeness Heuristic
- Schizophrenia
- Scientific Reasoning
- Semantic Dementia
- Spatial Cognition, Development of
- Thinking
- Two System Models of Reasoning
- Visual Imagery
- Visuospatial Reasoning
- Williams Syndrome
- Practical Applications
- Affective Forecasting
- Anxiety Disorders
- Attention and Emotion
- Cognitive Dissonance
- Descriptions
- Descriptive Thought
- Disgust
- Embarrassment
- Emotion and Moral Judgment
- Emotion and Working Memory
- Emotion Regulation
- Emotion, Psychophysiology of
- Emotion, Structural Approaches
- Envy
- Exercise and the Brain
- Facial Expressions, Emotional
- Guilt
- Happiness
- Indexical Thought
- Intension and Extension
- Jealousy
- Love
- Metaphor
- Mnemonic Strategies
- Object-Dependent Thought
- Regret
- Religion and Psychiatry
- Resentment
- Retrieval Practice (Testing) Effect
- Spacing Effect, Practical Applications
- Unconscious Emotions, Psychological Perspectives
- Psychological Research
- Aging, Memory, and Information Processing Speed
- Deception, Linguistic Cues to
- Divided Attention and Memory
- Emotion and Working Memory
- Event Memory, Development
- Exercise and the Brain
- Eyewitness Memory
- Implicit Memory
- Intelligence and Working Memory
- Lie Detection
- Memory Recall, Dynamics
- Memory, Interference With
- Rehearsal and Memory
- Retrieval Practice (Testing) Effect
- Semantic Memory
- Spacing Effect
- Visual Working Memory
- Working Memory
- Working Memory in Language Processing
- Philosophical Perspectives
- Action and Bodily Movement
- Analogical Reasoning, Models of Development
- Anomalous Monism
- Anomalous Monism
- Anti-Individualism About Cognition
- Anti-Individualism About Cognition
- Attention and Emotions, Computational Perspectives
- Belief and Judgment
- Case-Based Reasoning, Computational Perspectives
- Causal Theories of Intentionality
- Causal Theories of Memory
- Computational Models of Emotion
- Computational Perspectives
- Conscious Thinking
- Conscious Thinking
- Consciousness and Embodiment
- Consciousness and Embodiment
- Consciousness and the Unconscious
- Deductive Reasoning
- Descriptive Thought
- Development
- Disorders and Pathology
- Disorders and Pathology
- Distributed Cognition
- Electrophysiological Studies of Mind
- Eliminative Materialism
- Eliminative Materialism
- Emergence
- Emergence
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Experimental Philosophy
- Explanatory Gap
- Explanatory Gap
- Extended Mind
- Face Recognition in Humans and Computers
- Facial Expressions, Computational Perspectives
- Facial Expressions, Computational Perspectives
- Freedom of Action
- Idealism
- Idealism
- Intelligence, Neural Basis
- Intentionality of Bodily Sensation
- Intentionality of Emotion
- Introspection
- Know-How, Philosophical Perspectives
- Know-How, Philosophical Perspectives
- Knowledge by Acquaintance
- Machine Speech Recognition
- Memory and Knowledge
- Mental Action
- Mental Causation
- Mental Causation
- Mind-Body Problem
- Modeling Causal Learning
- Moral Development
- Multimodal Conversational Systems
- Naïve Realism
- Natural Language Generation
- Neural Basis
- Object-Dependent Thought
- Personal Identity
- Personal Identity, Development of
- Philosophical Perspectives
- Philosophy of Action
- Physicalism
- Physicalism
- Psychological Research
- Realism and Instrumentalism
- Realism and Instrumentalism
- Reductive Physicalism
- Reductive Physicalism
- Relationships, Development of
- Representational Theory of Mind
- Self-Knowledge
- Semantic Memory, Computational Perspectives
- Sequential Memory, Computational Perspectives
- Serial Order Memory, Computational Perspectives
- Smell, Philosophical Perspectives
- Taste, Philosophical Perspectives
- Teleology
- Teleology
- Theory of Appearing
- Neural Basis
- Action and Bodily Movement
- Analogical Reasoning, Models of Development
- Anomalous Monism
- Anomalous Monism
- Anti-Individualism About Cognition
- Anti-Individualism About Cognition
- Attention and Emotions, Computational Perspectives
- Belief and Judgment
- Case-Based Reasoning, Computational Perspectives
- Causal Theories of Intentionality
- Causal Theories of Memory
- Computational Models of Emotion
- Computational Perspectives
- Conscious Thinking
- Conscious Thinking
- Consciousness and Embodiment
- Consciousness and Embodiment
- Consciousness and the Unconscious
- Deductive Reasoning
- Descriptive Thought
- Development
- Disorders and Pathology
- Disorders and Pathology
- Distributed Cognition
- Electrophysiological Studies of Mind
- Eliminative Materialism
- Eliminative Materialism
- Emergence
- Emergence
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Experimental Philosophy
- Explanatory Gap
- Explanatory Gap
- Extended Mind
- Face Recognition in Humans and Computers
- Facial Expressions, Computational Perspectives
- Facial Expressions, Computational Perspectives
- Freedom of Action
- Idealism
- Idealism
- Intelligence, Neural Basis
- Intentionality of Bodily Sensation
- Intentionality of Emotion
- Introspection
- Know-How, Philosophical Perspectives
- Know-How, Philosophical Perspectives
- Knowledge by Acquaintance
- Machine Speech Recognition
- Memory and Knowledge
- Mental Action
- Mental Causation
- Mental Causation
- Mind-Body Problem
- Modeling Causal Learning
- Moral Development
- Multimodal Conversational Systems
- Naïve Realism
- Natural Language Generation
- Neural Basis
- Object-Dependent Thought
- Personal Identity
- Personal Identity, Development of
- Philosophical Perspectives
- Philosophy of Action
- Physicalism
- Physicalism
- Psychological Research
- Realism and Instrumentalism
- Realism and Instrumentalism
- Reductive Physicalism
- Reductive Physicalism
- Relationships, Development of
- Representational Theory of Mind
- Self-Knowledge
- Semantic Memory, Computational Perspectives
- Sequential Memory, Computational Perspectives
- Serial Order Memory, Computational Perspectives
- Smell, Philosophical Perspectives
- Taste, Philosophical Perspectives
- Teleology
- Teleology
- Theory of Appearing
- Philosophical Perspectives
- Aging, Memory, and Information Processing Speed
- Audition, Neural Basis
- Bilingual Language Processing
- Bilingualism, Cognitive Benefits of
- Black English Vernacular (Ebonics)
- Blindsight
- Borderline Personality Disorder
- Capgras Delusion
- Character and Personality, Philosophical Perspectives
- Collective Action
- Common Coding
- Computational Perspectives
- Distributed Cognition
- Emotion and Moral Judgment
- Emotion, Cultural Perspectives
- Emotional Recognition, Neuropsychology of
- Event Memory, Development
- Evolutionary Perspectives
- Experimental Philosophy
- Flynn Effect: Rising Intelligence Scores
- Gender Differences in Language and Language Use
- Gesture and Language Processing
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Heritability
- Heritage Language and Second Language Learning
- Intelligence and Working Memory
- Joint or Collective Intention
- Joint or Collective Intention
- Knowledge Acquisition in Development
- Mirror Neurons
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Multiple Intelligences Theory
- Narcissistic Personality Disorder
- Neural Basis
- Philosophical Perspectives
- Philosophy of Action
- Psychological Research
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Self-Knowledge
- Synesthesia
- Visual Imagery
- Word Learning
- Practical Applications
- Amnesia
- Analogical Reasoning, Models of Development
- Anxiety Disorders
- Character and Personality, Philosophical Perspectives
- Computational Perspectives
- Confabulation
- Desire
- Development
- Disjunctive Theory of Perception
- Disorders and Pathology
- Dissent, Effects on Group Decisions
- Distributed Cognition
- Dyslexia, Developmental
- Dyslexia, Phonological Processing in
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Exercise and the Brain
- Folk Psychology
- Group Decision Making
- Guilt
- Hearing, Philosophical Perspectives
- Heritage Language and Second Language Learning
- Human Classification Learning
- Innateness and Parameter Setting
- Intergroup Conflict
- Intergroup Conflict, Models of
- Knowledge Acquisition in Development
- Language Development
- Language Development, Overregulation in
- Moral Development
- Naïve Realism
- Neural Basis
- Philosophical Perspectives
- Political Psychology
- Prediction, Clinical Versus Actuarial
- Psychological Research
- Religion and Psychiatry
- Semantic Dementia
- Smell, Philosophical Perspectives
- Spatial Cognition, Development of
- Taste, Philosophical Perspectives
- Word Learning
- Psychological Research
- Aphasia
- Attitude Change
- Attitude Change
- Attitudes and Behavior
- Attraction
- Attribution Theory
- Auditory Masking
- Behavioral Therapy
- Borderline Personality Disorder
- Cognitive Dissonance
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Common Coding
- Conduction Aphasia
- Consciousness, Comparative Perspectives
- Deductive Reasoning
- Depth Perception
- Desirable Difficulties Perspective on Learning
- Discrimination Learning, Training Methods
- Disorders and Pathology
- Dyslexia, Acquired
- Dyslexia, Developmental
- Dyslexia, Phonological Processing in
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Evolutionary Perspectives
- Exercise and the Brain
- Face Perception
- Facial Expressions, Emotional
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Group Decision Making
- Hearing
- Heritability
- Intergroup Conflict
- Intergroup Conflict, Models of
- Learning Styles
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Love
- McCollough Effect
- Meta-Analysis
- Metacognition and Education
- Motor Learning, Practical Aspects
- Multinomial Modeling
- Music Perception
- Neural Basis
- Optic Flow
- Perceptual Consciousness and Attention
- Perceptual Constancy
- Personal Identity, Development of
- Personality: Individual Versus Situation Debate
- Personality: Individual Versus Situation Debate
- Persuasion
- Philosophical Perspectives
- Placebo Effect
- Political Psychology
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Political Psychology
- Psychological Research
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Reaction Time
- Retrieval Practice (Testing) Effect
- Semantic Dementia
- Skill Learning, Enhancement of
- Social Cognition
- Social Loafing
- Spacing Effect, Practical Applications
- Speech Perception
- Stereopsis
- Subliminal Perception
- Synesthesia
- Time Perception
- Unconscious Perception
- Visual Imagery
- Visual Masking
- Visual Search
- Visuospatial Reasoning
- Wisdom of Crowds Effect
- Word Recognition, Auditory
- Word Recognition, Visual
- Working Memory, Evolution of
- Philosophical Perspectives
- Action and Bodily Movement
- Analogical Reasoning, Models of Development
- Anomalous Monism
- Anomalous Monism
- Anti-Individualism About Cognition
- Anti-Individualism About Cognition
- Attention and Emotions, Computational Perspectives
- Belief and Judgment
- Case-Based Reasoning, Computational Perspectives
- Causal Theories of Intentionality
- Causal Theories of Memory
- Computational Models of Emotion
- Computational Perspectives
- Conscious Thinking
- Conscious Thinking
- Consciousness and Embodiment
- Consciousness and Embodiment
- Consciousness and the Unconscious
- Deductive Reasoning
- Descriptive Thought
- Development
- Disorders and Pathology
- Disorders and Pathology
- Distributed Cognition
- Electrophysiological Studies of Mind
- Eliminative Materialism
- Eliminative Materialism
- Emergence
- Emergence
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Experimental Philosophy
- Explanatory Gap
- Explanatory Gap
- Extended Mind
- Face Recognition in Humans and Computers
- Facial Expressions, Computational Perspectives
- Facial Expressions, Computational Perspectives
- Freedom of Action
- Idealism
- Idealism
- Intelligence, Neural Basis
- Intentionality of Bodily Sensation
- Intentionality of Emotion
- Introspection
- Know-How, Philosophical Perspectives
- Know-How, Philosophical Perspectives
- Knowledge by Acquaintance
- Machine Speech Recognition
- Memory and Knowledge
- Mental Action
- Mental Causation
- Mental Causation
- Mind-Body Problem
- Modeling Causal Learning
- Moral Development
- Multimodal Conversational Systems
- Naïve Realism
- Natural Language Generation
- Neural Basis
- Object-Dependent Thought
- Personal Identity
- Personal Identity, Development of
- Philosophical Perspectives
- Philosophy of Action
- Physicalism
- Physicalism
- Psychological Research
- Realism and Instrumentalism
- Realism and Instrumentalism
- Reductive Physicalism
- Reductive Physicalism
- Relationships, Development of
- Representational Theory of Mind
- Self-Knowledge
- Semantic Memory, Computational Perspectives
- Sequential Memory, Computational Perspectives
- Serial Order Memory, Computational Perspectives
- Smell, Philosophical Perspectives
- Taste, Philosophical Perspectives
- Teleology
- Teleology
- Theory of Appearing
- Development
- Action and Bodily Movement
- Analogical Reasoning, Models of Development
- Anomalous Monism
- Anomalous Monism
- Anti-Individualism About Cognition
- Anti-Individualism About Cognition
- Attention and Emotions, Computational Perspectives
- Belief and Judgment
- Case-Based Reasoning, Computational Perspectives
- Causal Theories of Intentionality
- Causal Theories of Memory
- Computational Models of Emotion
- Computational Perspectives
- Conscious Thinking
- Conscious Thinking
- Consciousness and Embodiment
- Consciousness and Embodiment
- Consciousness and the Unconscious
- Deductive Reasoning
- Descriptive Thought
- Development
- Disorders and Pathology
- Disorders and Pathology
- Distributed Cognition
- Electrophysiological Studies of Mind
- Eliminative Materialism
- Eliminative Materialism
- Emergence
- Emergence
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Experimental Philosophy
- Explanatory Gap
- Explanatory Gap
- Extended Mind
- Face Recognition in Humans and Computers
- Facial Expressions, Computational Perspectives
- Facial Expressions, Computational Perspectives
- Freedom of Action
- Idealism
- Idealism
- Intelligence, Neural Basis
- Intentionality of Bodily Sensation
- Intentionality of Emotion
- Introspection
- Know-How, Philosophical Perspectives
- Know-How, Philosophical Perspectives
- Knowledge by Acquaintance
- Machine Speech Recognition
- Memory and Knowledge
- Mental Action
- Mental Causation
- Mental Causation
- Mind-Body Problem
- Modeling Causal Learning
- Moral Development
- Multimodal Conversational Systems
- Naïve Realism
- Natural Language Generation
- Neural Basis
- Object-Dependent Thought
- Personal Identity
- Personal Identity, Development of
- Philosophical Perspectives
- Philosophy of Action
- Physicalism
- Physicalism
- Psychological Research
- Realism and Instrumentalism
- Realism and Instrumentalism
- Reductive Physicalism
- Reductive Physicalism
- Relationships, Development of
- Representational Theory of Mind
- Self-Knowledge
- Semantic Memory, Computational Perspectives
- Sequential Memory, Computational Perspectives
- Serial Order Memory, Computational Perspectives
- Smell, Philosophical Perspectives
- Taste, Philosophical Perspectives
- Teleology
- Teleology
- Theory of Appearing
- Philosophical Perspectives
- Aging, Memory, and Information Processing Speed
- Audition, Neural Basis
- Bilingual Language Processing
- Bilingualism, Cognitive Benefits of
- Black English Vernacular (Ebonics)
- Blindsight
- Borderline Personality Disorder
- Capgras Delusion
- Character and Personality, Philosophical Perspectives
- Collective Action
- Common Coding
- Computational Perspectives
- Distributed Cognition
- Emotion and Moral Judgment
- Emotion, Cultural Perspectives
- Emotional Recognition, Neuropsychology of
- Event Memory, Development
- Evolutionary Perspectives
- Experimental Philosophy
- Flynn Effect: Rising Intelligence Scores
- Gender Differences in Language and Language Use
- Gesture and Language Processing
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Heritability
- Heritage Language and Second Language Learning
- Intelligence and Working Memory
- Joint or Collective Intention
- Joint or Collective Intention
- Knowledge Acquisition in Development
- Mirror Neurons
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Multiple Intelligences Theory
- Narcissistic Personality Disorder
- Neural Basis
- Philosophical Perspectives
- Philosophy of Action
- Psychological Research
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Self-Knowledge
- Synesthesia
- Visual Imagery
- Word Learning
- Psychological Research
- Amnesia
- Analogical Reasoning, Models of Development
- Anxiety Disorders
- Character and Personality, Philosophical Perspectives
- Computational Perspectives
- Confabulation
- Desire
- Development
- Disjunctive Theory of Perception
- Disorders and Pathology
- Dissent, Effects on Group Decisions
- Distributed Cognition
- Dyslexia, Developmental
- Dyslexia, Phonological Processing in
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Exercise and the Brain
- Folk Psychology
- Group Decision Making
- Guilt
- Hearing, Philosophical Perspectives
- Heritage Language and Second Language Learning
- Human Classification Learning
- Innateness and Parameter Setting
- Intergroup Conflict
- Intergroup Conflict, Models of
- Knowledge Acquisition in Development
- Language Development
- Language Development, Overregulation in
- Moral Development
- Naïve Realism
- Neural Basis
- Philosophical Perspectives
- Political Psychology
- Prediction, Clinical Versus Actuarial
- Psychological Research
- Religion and Psychiatry
- Semantic Dementia
- Smell, Philosophical Perspectives
- Spatial Cognition, Development of
- Taste, Philosophical Perspectives
- Word Learning
- Computational Perspectives
- Action and Bodily Movement
- Analogical Reasoning, Models of Development
- Anomalous Monism
- Anomalous Monism
- Anti-Individualism About Cognition
- Anti-Individualism About Cognition
- Attention and Emotions, Computational Perspectives
- Belief and Judgment
- Case-Based Reasoning, Computational Perspectives
- Causal Theories of Intentionality
- Causal Theories of Memory
- Computational Models of Emotion
- Computational Perspectives
- Conscious Thinking
- Conscious Thinking
- Consciousness and Embodiment
- Consciousness and Embodiment
- Consciousness and the Unconscious
- Deductive Reasoning
- Descriptive Thought
- Development
- Disorders and Pathology
- Disorders and Pathology
- Distributed Cognition
- Electrophysiological Studies of Mind
- Eliminative Materialism
- Eliminative Materialism
- Emergence
- Emergence
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Experimental Philosophy
- Explanatory Gap
- Explanatory Gap
- Extended Mind
- Face Recognition in Humans and Computers
- Facial Expressions, Computational Perspectives
- Facial Expressions, Computational Perspectives
- Freedom of Action
- Idealism
- Idealism
- Intelligence, Neural Basis
- Intentionality of Bodily Sensation
- Intentionality of Emotion
- Introspection
- Know-How, Philosophical Perspectives
- Know-How, Philosophical Perspectives
- Knowledge by Acquaintance
- Machine Speech Recognition
- Memory and Knowledge
- Mental Action
- Mental Causation
- Mental Causation
- Mind-Body Problem
- Modeling Causal Learning
- Moral Development
- Multimodal Conversational Systems
- Naïve Realism
- Natural Language Generation
- Neural Basis
- Object-Dependent Thought
- Personal Identity
- Personal Identity, Development of
- Philosophical Perspectives
- Philosophy of Action
- Physicalism
- Physicalism
- Psychological Research
- Realism and Instrumentalism
- Realism and Instrumentalism
- Reductive Physicalism
- Reductive Physicalism
- Relationships, Development of
- Representational Theory of Mind
- Self-Knowledge
- Semantic Memory, Computational Perspectives
- Sequential Memory, Computational Perspectives
- Serial Order Memory, Computational Perspectives
- Smell, Philosophical Perspectives
- Taste, Philosophical Perspectives
- Teleology
- Teleology
- Theory of Appearing
- Neural Basis
- Aging, Memory, and Information Processing Speed
- Audition, Neural Basis
- Bilingual Language Processing
- Bilingualism, Cognitive Benefits of
- Black English Vernacular (Ebonics)
- Blindsight
- Borderline Personality Disorder
- Capgras Delusion
- Character and Personality, Philosophical Perspectives
- Collective Action
- Common Coding
- Computational Perspectives
- Distributed Cognition
- Emotion and Moral Judgment
- Emotion, Cultural Perspectives
- Emotional Recognition, Neuropsychology of
- Event Memory, Development
- Evolutionary Perspectives
- Experimental Philosophy
- Flynn Effect: Rising Intelligence Scores
- Gender Differences in Language and Language Use
- Gesture and Language Processing
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Heritability
- Heritage Language and Second Language Learning
- Intelligence and Working Memory
- Joint or Collective Intention
- Joint or Collective Intention
- Knowledge Acquisition in Development
- Mirror Neurons
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Multiple Intelligences Theory
- Narcissistic Personality Disorder
- Neural Basis
- Philosophical Perspectives
- Philosophy of Action
- Psychological Research
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Self-Knowledge
- Synesthesia
- Visual Imagery
- Word Learning
- Philosophical Perspectives
- Amnesia
- Analogical Reasoning, Models of Development
- Anxiety Disorders
- Character and Personality, Philosophical Perspectives
- Computational Perspectives
- Confabulation
- Desire
- Development
- Disjunctive Theory of Perception
- Disorders and Pathology
- Dissent, Effects on Group Decisions
- Distributed Cognition
- Dyslexia, Developmental
- Dyslexia, Phonological Processing in
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Exercise and the Brain
- Folk Psychology
- Group Decision Making
- Guilt
- Hearing, Philosophical Perspectives
- Heritage Language and Second Language Learning
- Human Classification Learning
- Innateness and Parameter Setting
- Intergroup Conflict
- Intergroup Conflict, Models of
- Knowledge Acquisition in Development
- Language Development
- Language Development, Overregulation in
- Moral Development
- Naïve Realism
- Neural Basis
- Philosophical Perspectives
- Political Psychology
- Prediction, Clinical Versus Actuarial
- Psychological Research
- Religion and Psychiatry
- Semantic Dementia
- Smell, Philosophical Perspectives
- Spatial Cognition, Development of
- Taste, Philosophical Perspectives
- Word Learning
- Psychological Research
- Aphasia
- Attitude Change
- Attitude Change
- Attitudes and Behavior
- Attraction
- Attribution Theory
- Auditory Masking
- Behavioral Therapy
- Borderline Personality Disorder
- Cognitive Dissonance
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Common Coding
- Conduction Aphasia
- Consciousness, Comparative Perspectives
- Deductive Reasoning
- Depth Perception
- Desirable Difficulties Perspective on Learning
- Discrimination Learning, Training Methods
- Disorders and Pathology
- Dyslexia, Acquired
- Dyslexia, Developmental
- Dyslexia, Phonological Processing in
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Evolutionary Perspectives
- Exercise and the Brain
- Face Perception
- Facial Expressions, Emotional
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Group Decision Making
- Hearing
- Heritability
- Intergroup Conflict
- Intergroup Conflict, Models of
- Learning Styles
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Love
- McCollough Effect
- Meta-Analysis
- Metacognition and Education
- Motor Learning, Practical Aspects
- Multinomial Modeling
- Music Perception
- Neural Basis
- Optic Flow
- Perceptual Consciousness and Attention
- Perceptual Constancy
- Personal Identity, Development of
- Personality: Individual Versus Situation Debate
- Personality: Individual Versus Situation Debate
- Persuasion
- Philosophical Perspectives
- Placebo Effect
- Political Psychology
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Political Psychology
- Psychological Research
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Reaction Time
- Retrieval Practice (Testing) Effect
- Semantic Dementia
- Skill Learning, Enhancement of
- Social Cognition
- Social Loafing
- Spacing Effect, Practical Applications
- Speech Perception
- Stereopsis
- Subliminal Perception
- Synesthesia
- Time Perception
- Unconscious Perception
- Visual Imagery
- Visual Masking
- Visual Search
- Visuospatial Reasoning
- Wisdom of Crowds Effect
- Word Recognition, Auditory
- Word Recognition, Visual
- Working Memory, Evolution of
- Development
- Action and Bodily Movement
- Analogical Reasoning, Models of Development
- Anomalous Monism
- Anomalous Monism
- Anti-Individualism About Cognition
- Anti-Individualism About Cognition
- Attention and Emotions, Computational Perspectives
- Belief and Judgment
- Case-Based Reasoning, Computational Perspectives
- Causal Theories of Intentionality
- Causal Theories of Memory
- Computational Models of Emotion
- Computational Perspectives
- Conscious Thinking
- Conscious Thinking
- Consciousness and Embodiment
- Consciousness and Embodiment
- Consciousness and the Unconscious
- Deductive Reasoning
- Descriptive Thought
- Development
- Disorders and Pathology
- Disorders and Pathology
- Distributed Cognition
- Electrophysiological Studies of Mind
- Eliminative Materialism
- Eliminative Materialism
- Emergence
- Emergence
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Experimental Philosophy
- Explanatory Gap
- Explanatory Gap
- Extended Mind
- Face Recognition in Humans and Computers
- Facial Expressions, Computational Perspectives
- Facial Expressions, Computational Perspectives
- Freedom of Action
- Idealism
- Idealism
- Intelligence, Neural Basis
- Intentionality of Bodily Sensation
- Intentionality of Emotion
- Introspection
- Know-How, Philosophical Perspectives
- Know-How, Philosophical Perspectives
- Knowledge by Acquaintance
- Machine Speech Recognition
- Memory and Knowledge
- Mental Action
- Mental Causation
- Mental Causation
- Mind-Body Problem
- Modeling Causal Learning
- Moral Development
- Multimodal Conversational Systems
- Naïve Realism
- Natural Language Generation
- Neural Basis
- Object-Dependent Thought
- Personal Identity
- Personal Identity, Development of
- Philosophical Perspectives
- Philosophy of Action
- Physicalism
- Physicalism
- Psychological Research
- Realism and Instrumentalism
- Realism and Instrumentalism
- Reductive Physicalism
- Reductive Physicalism
- Relationships, Development of
- Representational Theory of Mind
- Self-Knowledge
- Semantic Memory, Computational Perspectives
- Sequential Memory, Computational Perspectives
- Serial Order Memory, Computational Perspectives
- Smell, Philosophical Perspectives
- Taste, Philosophical Perspectives
- Teleology
- Teleology
- Theory of Appearing
- Disorders and Pathology
- Aging, Memory, and Information Processing Speed
- Audition, Neural Basis
- Bilingual Language Processing
- Bilingualism, Cognitive Benefits of
- Black English Vernacular (Ebonics)
- Blindsight
- Borderline Personality Disorder
- Capgras Delusion
- Character and Personality, Philosophical Perspectives
- Collective Action
- Common Coding
- Computational Perspectives
- Distributed Cognition
- Emotion and Moral Judgment
- Emotion, Cultural Perspectives
- Emotional Recognition, Neuropsychology of
- Event Memory, Development
- Evolutionary Perspectives
- Experimental Philosophy
- Flynn Effect: Rising Intelligence Scores
- Gender Differences in Language and Language Use
- Gesture and Language Processing
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Heritability
- Heritage Language and Second Language Learning
- Intelligence and Working Memory
- Joint or Collective Intention
- Joint or Collective Intention
- Knowledge Acquisition in Development
- Mirror Neurons
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Multiple Intelligences Theory
- Narcissistic Personality Disorder
- Neural Basis
- Philosophical Perspectives
- Philosophy of Action
- Psychological Research
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Self-Knowledge
- Synesthesia
- Visual Imagery
- Word Learning
- Philosophical Perspectives
- Amnesia
- Analogical Reasoning, Models of Development
- Anxiety Disorders
- Character and Personality, Philosophical Perspectives
- Computational Perspectives
- Confabulation
- Desire
- Development
- Disjunctive Theory of Perception
- Disorders and Pathology
- Dissent, Effects on Group Decisions
- Distributed Cognition
- Dyslexia, Developmental
- Dyslexia, Phonological Processing in
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Exercise and the Brain
- Folk Psychology
- Group Decision Making
- Guilt
- Hearing, Philosophical Perspectives
- Heritage Language and Second Language Learning
- Human Classification Learning
- Innateness and Parameter Setting
- Intergroup Conflict
- Intergroup Conflict, Models of
- Knowledge Acquisition in Development
- Language Development
- Language Development, Overregulation in
- Moral Development
- Naïve Realism
- Neural Basis
- Philosophical Perspectives
- Political Psychology
- Prediction, Clinical Versus Actuarial
- Psychological Research
- Religion and Psychiatry
- Semantic Dementia
- Smell, Philosophical Perspectives
- Spatial Cognition, Development of
- Taste, Philosophical Perspectives
- Word Learning
- Psychological Research
- Aphasia
- Attitude Change
- Attitude Change
- Attitudes and Behavior
- Attraction
- Attribution Theory
- Auditory Masking
- Behavioral Therapy
- Borderline Personality Disorder
- Cognitive Dissonance
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Common Coding
- Conduction Aphasia
- Consciousness, Comparative Perspectives
- Deductive Reasoning
- Depth Perception
- Desirable Difficulties Perspective on Learning
- Discrimination Learning, Training Methods
- Disorders and Pathology
- Dyslexia, Acquired
- Dyslexia, Developmental
- Dyslexia, Phonological Processing in
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Evolutionary Perspectives
- Exercise and the Brain
- Face Perception
- Facial Expressions, Emotional
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Group Decision Making
- Hearing
- Heritability
- Intergroup Conflict
- Intergroup Conflict, Models of
- Learning Styles
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Love
- McCollough Effect
- Meta-Analysis
- Metacognition and Education
- Motor Learning, Practical Aspects
- Multinomial Modeling
- Music Perception
- Neural Basis
- Optic Flow
- Perceptual Consciousness and Attention
- Perceptual Constancy
- Personal Identity, Development of
- Personality: Individual Versus Situation Debate
- Personality: Individual Versus Situation Debate
- Persuasion
- Philosophical Perspectives
- Placebo Effect
- Political Psychology
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Political Psychology
- Psychological Research
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Reaction Time
- Retrieval Practice (Testing) Effect
- Semantic Dementia
- Skill Learning, Enhancement of
- Social Cognition
- Social Loafing
- Spacing Effect, Practical Applications
- Speech Perception
- Stereopsis
- Subliminal Perception
- Synesthesia
- Time Perception
- Unconscious Perception
- Visual Imagery
- Visual Masking
- Visual Search
- Visuospatial Reasoning
- Wisdom of Crowds Effect
- Word Recognition, Auditory
- Word Recognition, Visual
- Working Memory, Evolution of
- Development
- Action and Bodily Movement
- Analogical Reasoning, Models of Development
- Anomalous Monism
- Anomalous Monism
- Anti-Individualism About Cognition
- Anti-Individualism About Cognition
- Attention and Emotions, Computational Perspectives
- Belief and Judgment
- Case-Based Reasoning, Computational Perspectives
- Causal Theories of Intentionality
- Causal Theories of Memory
- Computational Models of Emotion
- Computational Perspectives
- Conscious Thinking
- Conscious Thinking
- Consciousness and Embodiment
- Consciousness and Embodiment
- Consciousness and the Unconscious
- Deductive Reasoning
- Descriptive Thought
- Development
- Disorders and Pathology
- Disorders and Pathology
- Distributed Cognition
- Electrophysiological Studies of Mind
- Eliminative Materialism
- Eliminative Materialism
- Emergence
- Emergence
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Experimental Philosophy
- Explanatory Gap
- Explanatory Gap
- Extended Mind
- Face Recognition in Humans and Computers
- Facial Expressions, Computational Perspectives
- Facial Expressions, Computational Perspectives
- Freedom of Action
- Idealism
- Idealism
- Intelligence, Neural Basis
- Intentionality of Bodily Sensation
- Intentionality of Emotion
- Introspection
- Know-How, Philosophical Perspectives
- Know-How, Philosophical Perspectives
- Knowledge by Acquaintance
- Machine Speech Recognition
- Memory and Knowledge
- Mental Action
- Mental Causation
- Mental Causation
- Mind-Body Problem
- Modeling Causal Learning
- Moral Development
- Multimodal Conversational Systems
- Naïve Realism
- Natural Language Generation
- Neural Basis
- Object-Dependent Thought
- Personal Identity
- Personal Identity, Development of
- Philosophical Perspectives
- Philosophy of Action
- Physicalism
- Physicalism
- Psychological Research
- Realism and Instrumentalism
- Realism and Instrumentalism
- Reductive Physicalism
- Reductive Physicalism
- Relationships, Development of
- Representational Theory of Mind
- Self-Knowledge
- Semantic Memory, Computational Perspectives
- Sequential Memory, Computational Perspectives
- Serial Order Memory, Computational Perspectives
- Smell, Philosophical Perspectives
- Taste, Philosophical Perspectives
- Teleology
- Teleology
- Theory of Appearing
- Disorders and Pathology
- Aging, Memory, and Information Processing Speed
- Audition, Neural Basis
- Bilingual Language Processing
- Bilingualism, Cognitive Benefits of
- Black English Vernacular (Ebonics)
- Blindsight
- Borderline Personality Disorder
- Capgras Delusion
- Character and Personality, Philosophical Perspectives
- Collective Action
- Common Coding
- Computational Perspectives
- Distributed Cognition
- Emotion and Moral Judgment
- Emotion, Cultural Perspectives
- Emotional Recognition, Neuropsychology of
- Event Memory, Development
- Evolutionary Perspectives
- Experimental Philosophy
- Flynn Effect: Rising Intelligence Scores
- Gender Differences in Language and Language Use
- Gesture and Language Processing
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Heritability
- Heritage Language and Second Language Learning
- Intelligence and Working Memory
- Joint or Collective Intention
- Joint or Collective Intention
- Knowledge Acquisition in Development
- Mirror Neurons
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Multiple Intelligences Theory
- Narcissistic Personality Disorder
- Neural Basis
- Philosophical Perspectives
- Philosophy of Action
- Psychological Research
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Self-Knowledge
- Synesthesia
- Visual Imagery
- Word Learning
- Philosophical Perspectives
- Amnesia
- Analogical Reasoning, Models of Development
- Anxiety Disorders
- Character and Personality, Philosophical Perspectives
- Computational Perspectives
- Confabulation
- Desire
- Development
- Disjunctive Theory of Perception
- Disorders and Pathology
- Dissent, Effects on Group Decisions
- Distributed Cognition
- Dyslexia, Developmental
- Dyslexia, Phonological Processing in
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Exercise and the Brain
- Folk Psychology
- Group Decision Making
- Guilt
- Hearing, Philosophical Perspectives
- Heritage Language and Second Language Learning
- Human Classification Learning
- Innateness and Parameter Setting
- Intergroup Conflict
- Intergroup Conflict, Models of
- Knowledge Acquisition in Development
- Language Development
- Language Development, Overregulation in
- Moral Development
- Naïve Realism
- Neural Basis
- Philosophical Perspectives
- Political Psychology
- Prediction, Clinical Versus Actuarial
- Psychological Research
- Religion and Psychiatry
- Semantic Dementia
- Smell, Philosophical Perspectives
- Spatial Cognition, Development of
- Taste, Philosophical Perspectives
- Word Learning
- Psychological Research
- Aphasia
- Attitude Change
- Attitude Change
- Attitudes and Behavior
- Attraction
- Attribution Theory
- Auditory Masking
- Behavioral Therapy
- Borderline Personality Disorder
- Cognitive Dissonance
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Common Coding
- Conduction Aphasia
- Consciousness, Comparative Perspectives
- Deductive Reasoning
- Depth Perception
- Desirable Difficulties Perspective on Learning
- Discrimination Learning, Training Methods
- Disorders and Pathology
- Dyslexia, Acquired
- Dyslexia, Developmental
- Dyslexia, Phonological Processing in
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Evolutionary Perspectives
- Exercise and the Brain
- Face Perception
- Facial Expressions, Emotional
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Group Decision Making
- Hearing
- Heritability
- Intergroup Conflict
- Intergroup Conflict, Models of
- Learning Styles
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Love
- McCollough Effect
- Meta-Analysis
- Metacognition and Education
- Motor Learning, Practical Aspects
- Multinomial Modeling
- Music Perception
- Neural Basis
- Optic Flow
- Perceptual Consciousness and Attention
- Perceptual Constancy
- Personal Identity, Development of
- Personality: Individual Versus Situation Debate
- Personality: Individual Versus Situation Debate
- Persuasion
- Philosophical Perspectives
- Placebo Effect
- Political Psychology
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Political Psychology
- Psychological Research
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Reaction Time
- Retrieval Practice (Testing) Effect
- Semantic Dementia
- Skill Learning, Enhancement of
- Social Cognition
- Social Loafing
- Spacing Effect, Practical Applications
- Speech Perception
- Stereopsis
- Subliminal Perception
- Synesthesia
- Time Perception
- Unconscious Perception
- Visual Imagery
- Visual Masking
- Visual Search
- Visuospatial Reasoning
- Wisdom of Crowds Effect
- Word Recognition, Auditory
- Word Recognition, Visual
- Working Memory, Evolution of
- Computational Perspectives
- Action and Bodily Movement
- Analogical Reasoning, Models of Development
- Anomalous Monism
- Anomalous Monism
- Anti-Individualism About Cognition
- Anti-Individualism About Cognition
- Attention and Emotions, Computational Perspectives
- Belief and Judgment
- Case-Based Reasoning, Computational Perspectives
- Causal Theories of Intentionality
- Causal Theories of Memory
- Computational Models of Emotion
- Computational Perspectives
- Conscious Thinking
- Conscious Thinking
- Consciousness and Embodiment
- Consciousness and Embodiment
- Consciousness and the Unconscious
- Deductive Reasoning
- Descriptive Thought
- Development
- Disorders and Pathology
- Disorders and Pathology
- Distributed Cognition
- Electrophysiological Studies of Mind
- Eliminative Materialism
- Eliminative Materialism
- Emergence
- Emergence
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Experimental Philosophy
- Explanatory Gap
- Explanatory Gap
- Extended Mind
- Face Recognition in Humans and Computers
- Facial Expressions, Computational Perspectives
- Facial Expressions, Computational Perspectives
- Freedom of Action
- Idealism
- Idealism
- Intelligence, Neural Basis
- Intentionality of Bodily Sensation
- Intentionality of Emotion
- Introspection
- Know-How, Philosophical Perspectives
- Know-How, Philosophical Perspectives
- Knowledge by Acquaintance
- Machine Speech Recognition
- Memory and Knowledge
- Mental Action
- Mental Causation
- Mental Causation
- Mind-Body Problem
- Modeling Causal Learning
- Moral Development
- Multimodal Conversational Systems
- Naïve Realism
- Natural Language Generation
- Neural Basis
- Object-Dependent Thought
- Personal Identity
- Personal Identity, Development of
- Philosophical Perspectives
- Philosophy of Action
- Physicalism
- Physicalism
- Psychological Research
- Realism and Instrumentalism
- Realism and Instrumentalism
- Reductive Physicalism
- Reductive Physicalism
- Relationships, Development of
- Representational Theory of Mind
- Self-Knowledge
- Semantic Memory, Computational Perspectives
- Sequential Memory, Computational Perspectives
- Serial Order Memory, Computational Perspectives
- Smell, Philosophical Perspectives
- Taste, Philosophical Perspectives
- Teleology
- Teleology
- Theory of Appearing
- Cultural Perspectives
- Aging, Memory, and Information Processing Speed
- Audition, Neural Basis
- Bilingual Language Processing
- Bilingualism, Cognitive Benefits of
- Black English Vernacular (Ebonics)
- Blindsight
- Borderline Personality Disorder
- Capgras Delusion
- Character and Personality, Philosophical Perspectives
- Collective Action
- Common Coding
- Computational Perspectives
- Distributed Cognition
- Emotion and Moral Judgment
- Emotion, Cultural Perspectives
- Emotional Recognition, Neuropsychology of
- Event Memory, Development
- Evolutionary Perspectives
- Experimental Philosophy
- Flynn Effect: Rising Intelligence Scores
- Gender Differences in Language and Language Use
- Gesture and Language Processing
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Heritability
- Heritage Language and Second Language Learning
- Intelligence and Working Memory
- Joint or Collective Intention
- Joint or Collective Intention
- Knowledge Acquisition in Development
- Mirror Neurons
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Multiple Intelligences Theory
- Narcissistic Personality Disorder
- Neural Basis
- Philosophical Perspectives
- Philosophy of Action
- Psychological Research
- Attention and Action
- Attention and Emotion
- Attention, Resource Models
- Attentional Blink Effect
- Automaticity
- Change Blindness
- Divided Attention and Memory
- Inattentional Blindness
- Inhibition of Return
- Mental Effort
- Multitasking and Human Performance
- Neurodynamics of Visual Search
- Perceptual Consciousness and Attention
- Psychological Refractory Period
- Stroop Effect
- Visual Search
- Self-Knowledge
- Synesthesia
- Visual Imagery
- Word Learning
- Development
- Amnesia
- Analogical Reasoning, Models of Development
- Anxiety Disorders
- Character and Personality, Philosophical Perspectives
- Computational Perspectives
- Confabulation
- Desire
- Development
- Disjunctive Theory of Perception
- Disorders and Pathology
- Dissent, Effects on Group Decisions
- Distributed Cognition
- Dyslexia, Developmental
- Dyslexia, Phonological Processing in
- Emotion and Moral Judgment
- Evolutionary Perspectives
- Exercise and the Brain
- Folk Psychology
- Group Decision Making
- Guilt
- Hearing, Philosophical Perspectives
- Heritage Language and Second Language Learning
- Human Classification Learning
- Innateness and Parameter Setting
- Intergroup Conflict
- Intergroup Conflict, Models of
- Knowledge Acquisition in Development
- Language Development
- Language Development, Overregulation in
- Moral Development
- Naïve Realism
- Neural Basis
- Philosophical Perspectives
- Political Psychology
- Prediction, Clinical Versus Actuarial
- Psychological Research
- Religion and Psychiatry
- Semantic Dementia
- Smell, Philosophical Perspectives
- Spatial Cognition, Development of
- Taste, Philosophical Perspectives
- Word Learning
- Philosophical Perspectives
- Aphasia
- Attitude Change
- Attitude Change
- Attitudes and Behavior
- Attraction
- Attribution Theory
- Auditory Masking
- Behavioral Therapy
- Borderline Personality Disorder
- Cognitive Dissonance
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Common Coding
- Conduction Aphasia
- Consciousness, Comparative Perspectives
- Deductive Reasoning
- Depth Perception
- Desirable Difficulties Perspective on Learning
- Discrimination Learning, Training Methods
- Disorders and Pathology
- Dyslexia, Acquired
- Dyslexia, Developmental
- Dyslexia, Phonological Processing in
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Evolutionary Perspectives
- Exercise and the Brain
- Face Perception
- Facial Expressions, Emotional
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Group Decision Making
- Hearing
- Heritability
- Intergroup Conflict
- Intergroup Conflict, Models of
- Learning Styles
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Love
- McCollough Effect
- Meta-Analysis
- Metacognition and Education
- Motor Learning, Practical Aspects
- Multinomial Modeling
- Music Perception
- Neural Basis
- Optic Flow
- Perceptual Consciousness and Attention
- Perceptual Constancy
- Personal Identity, Development of
- Personality: Individual Versus Situation Debate
- Personality: Individual Versus Situation Debate
- Persuasion
- Philosophical Perspectives
- Placebo Effect
- Political Psychology
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Political Psychology
- Psychological Research
- Anesthesia and Awareness
- Attention and Consciousness
- Consciousness and the Unconscious
- Metacognition and Education
- Perceptual Consciousness and Attention
- Self-Consciousness
- Self-Knowledge
- Subliminal Perception
- Unconscious Emotions, Psychological Perspectives
- Unconscious Perception
- Voluntary Action, Illusion of
- Reaction Time
- Retrieval Practice (Testing) Effect
- Semantic Dementia
- Skill Learning, Enhancement of
- Social Cognition
- Social Loafing
- Spacing Effect, Practical Applications
- Speech Perception
- Stereopsis
- Subliminal Perception
- Synesthesia
- Time Perception
- Unconscious Perception
- Visual Imagery
- Visual Masking
- Visual Search
- Visuospatial Reasoning
- Wisdom of Crowds Effect
- Word Recognition, Auditory
- Word Recognition, Visual
- Working Memory, Evolution of
- Practical Applications
- Amnesia
- Argument Mapping
- Behavioral Therapy
- Behaviorism
- Case-Based Reasoning, Computational Perspectives
- Computational Perspectives
- Desirable Difficulties Perspective on Learning
- Desire
- Desire
- Emotion and Psychopathology
- Emotion, Cerebral Lateralization
- Emotion, Psychophysiology of
- Emotional Recognition, Neuropsychology of
- Event Memory, Development
- Genes and Linguistic Tone
- Human Classification Learning
- Implicit Memory
- Learning Styles
- Memory, Neural Basis
- Metacognition and Education
- Music and the Evolution of Language
- Neural Basis
- Philosophical Perspectives
- Placebo Effect
- Practical Applications
- Psychological Research
- Reinforcement Learning, Psychological Perspectives
- Retrieval Practice (Testing) Effect
- Skill Learning, Enhancement of
- Spacing Effect
- Word Learning
- Psychological Research
- Addiction
- Amnesia
- Analogical Mapping and Reasoning
- Anchoring
- Anosognosia
- Anxiety Disorders
- Aphasia
- Aphasia
- Apraxia
- Autism
- Automatic Behavior
- Autoscopic Phenomena
- Availability Heuristic
- Behavioral Therapy
- Borderline Personality Disorder
- Capgras Delusion
- Causal Theories of Memory
- Conduction Aphasia
- Conduction Aphasia
- Confabulation
- Delusions
- Desire
- Distributed Cognition
- Dyslexia, Acquired
- Dyslexia, Developmental
- Dyslexia, Phonological Processing in
- Emotion and Moral Judgment
- Emotion and Psychopathology
- Emotions and Consciousness
- Fregoli Delusion
- Hypochondria
- Intentionality of Emotion
- Legal Reasoning, Psychological Perspectives
- Memory and Knowledge
- Motivated Thinking
- Narcissistic Personality Disorder
- Objects of Memory
- Rationality of Emotion
- Religion and Psychiatry
- Representativeness Heuristic
- Schizophrenia
- Scientific Reasoning
- Semantic Dementia
- Spatial Cognition, Development of
- Thinking
- Two System Models of Reasoning
- Visual Imagery
- Visuospatial Reasoning
- Williams Syndrome
- Loading...
Get a 30 day FREE TRIAL
-
 Watch videos from a variety of sources bringing classroom topics to life
Watch videos from a variety of sources bringing classroom topics to life -
Read modern, diverse business cases
-
Explore hundreds of books and reference titles

Read next

More like this
Sage Recommends
We found other relevant content for you on other Sage platforms.
Have you created a personal profile? Login or create a profile so that you can save clips, playlists and searches